Studies of the adrenal medulla have provided several scientific "firsts." George Oliver and Edward Albert Schäfer were the first to extract a hormone from an endocrine organ when they showed that a preparation from the adrenal medulla had a profound effect on blood pressure. John Jacob Abel was the first to isolate a hormone when he isolated epinephrine from the adrenal medulla. Jokichi Takamine and Thomas Bell Aldrich, independently, were the first to crystallize a hormone, an important step on the way to chemical identification. Ernst Joseph Friedmann was the first to characterize a hormone by revealing its chemical formula. Hermann Blaschko was the first to outline the biosynthetic pathway of a hormone. Blaschko and Arnold Welch, and Nils-Åke Hillarp, Sten Lagerstedt and Bodil Nilson, were the first to isolate secretary vesicles, the chromaffin vesicle of adrenal medullary cells. This non-exhaustive list of studies with the adrenal medulla, and its principal hormone epinephrine, indicates that this organ has played an important and interesting role in the history of our understanding of the endocrine system and, as we shall see, the nervous system as well. But before we discuss these relatively recent breakthroughs, the events leading up to our current understanding of the adrenal medulla will be examined.
Since in the human, as in all mammals, the adrenal medulla is
completely enclosed by the adrenal cortex, the early history of
the medulla is necessarily linked to the discovery of the adrenal
glands. In fact, it was not recognized until relatively recently
that two functionally distinct organs coexisted within the adrenal
gland. By some accounts, the first description of the adrenal
glands is found in the Bible, specifically Leviticus 3:4. This
passage reads (in the King James version, and some key words are
different in other versions), "And the two kidneys, and the
fat that is on them, which is by the flanks......". The question
of whether or not the fat described in the passage refers to the
adrenal glands apparently comes down to technicalities of translation,
since the word for fat and the description of the relationship
of this fat to the kidney are questionable. However, at the most
generous, this cannot be considered a detailed description of
the adrenal glands.
Another controversy surrounds
the accuracy of the descriptions of the adrenal glands by Claudius
Galen (c. 130-201). From the German translation of Galen's seven
books by Simon in 190696, it is apparent that Galen encountered
the adrenal glands in his numerous dissections of animals. However,
only the left gland is described in his writings. While the gland
is described as "loose flesh," he clearly describes
the left adrenal vein connected to the left renal vein. Like many
anatomists in the centuries to follow, Galen assumed that this
"loose flesh" represented accessory renal tissue. Some
historians have not recognized the description of Galen as applying
specifically to the adrenal glands, but his accurate description
of the left adrenal vein leads one to the conclusion that Galen
was the first to publish a description of the mammalian adrenal
gland.
The first anatomist to give a detailed description of the human adrenal glands, including accurate illustrations, was Bartholomeus Eustachius34. Eustachius (1520- 1574) was professor at the Collegio della Sapienza at Rome. A significant part of his career was spent preparing detailed copper plates of the human anatomy. The
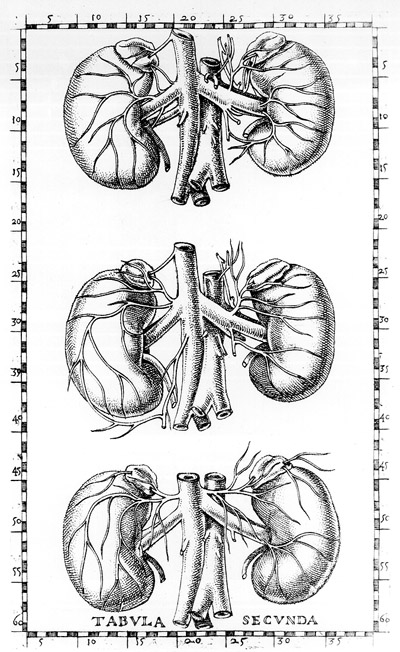
forty-seventh (and last)
of these splendid plates was completed in 1552. Interestingly,
these plates were placed in the Papal Library where they remained
for over a century. Rolleston89
reported that Pope
Clement XI gave them to his medical attendant, G.M. Lancisi, who
published them with his own text in 1714. Eustachius published
a description of the kidneys in 1563 and referred to the adrenal
glands as Glandulae Renibus Incumbentes, a name that implied
an accessory renal role. His description, here translated by Lenard65,
still holds true. "Even if many will consider sufficient
what we have said about the surface of the kidneys, somebody could
well object that I have neglected something and I consider it
indicated to say something of the glands, diligently overlooked
[diligenter praetermissis] by other anatomists. Both kidneys are
capped on the extremity towards the cava by a gland. Both are
connected with a fold of the peritoneum in such a way that one,
if he is not very attentive, does really overlook them, as if
they were not present. Their shape resembles that of the kidneys...sometimes
one is bigger, sometimes another...early anatomists and those
who write ample treatises on this art in our days failed to detect
them. They, pretending to be exact, stand so obstinately for their
own and their master's errors that often they seem to be fighters
rather than searchers of anatomical truth."
It is easy to see that
the politics of science flourished in Eustachius' day, perhaps
more openly than today. This bitter trend was to continue throughout
most of the history of the adrenal medulla. Lenard65 referred
to this era as that of the "fighting anatomists" [anatomici
contentiosi]. The next blow was delivered by Archangelo Piccolomini
who attacked Eustachius in 1586. An excerpt from his Anatomicae
prelectiones (from65): "Sometimes one may
see two or more glands lying on the kidneys but we do not think
they deserve special attention, because they are not to be found
in every case and they have no flesh or parenchyma of their own
... so that they might be considered renal excrescences. 'They
form part of the renal parenchyma. Why do they exist in a few?
The same way supra-abundance of material creates a sixth finger,
they originate from a seminal surplus and that is the way they
come out of the kidneys."
Some time later, in 1640,
André DuLaurens, physician of Henry IV of France, also
doubted the existence of the adrenal glands31.
He wrote: "Eustachius claims to find a gland above the
kidneys. Sometimes we saw that too; often, however, we stated
that there was no such gland."
Despite these, and other,
detractors, history shows that Eustachius was the first to describe
the adrenal glands accurately. Certainly part of the problem of
why this was not recognized by his contemporaries was due to the
suppression of his beautifully detailed copper plates. We will
probably never know exactly why this occurred, but one can speculate
that there were negative feelings, held by at least one influential
person under the Pope's authority, about the publication of illustrations
of human dissections. It apparently was not taboo for Eustachius
to publish a written description in 1563. Although the date of
1563 is often given as the earliest description of the adrenal
glands, the true date should be set as prior to 1552 when Eustachius
completed his plates.
As described by Goldzieher40,
other anatomists of the period, including Caspar Bauhin in 1588
and Archangelus Piccolhomineus in 1596, commented on the existence
of the adrenal glands. In 1611, another description of the adrenal
glands was put forward, which was to have a profound effect on
the contemporary concept of the adrenal medulla. The Danish anatomist
Caspar Bartholin (1585-1629), a student of Bauhin, described the
adrenal glands as hollow organs, filled with "black bile."
He gave them the name capsulae atrabiliarae. In his plates
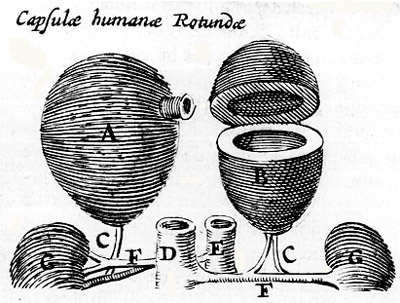 the spherical, hollow nature of
the glands is emphasized. Apparently, Bartholin's concept was
based on the erroneous description by Johann Schenk von Grafenburg
(1530-1598) published in 160090 of a central cavity in the gland,
corroborated and supplemented by Bauhin's "discovery,"
in 1605, of a black fluid in the cavity. This concept, and the
same plates, were passed on by Caspar's son, Thomas. Thomas Bartholin
(1616-1680)7, the second of six sons, was
destined to become more famous than his father. And it was Thomas'
son, Caspar the Younger, who described the greater vestibular
glands of the female perineum. An interesting history of the Bartholins,
who were well known as men of letters in addition to being famous
anatomists, is provided by Sir Humphry Davy Rolleston89.
the spherical, hollow nature of
the glands is emphasized. Apparently, Bartholin's concept was
based on the erroneous description by Johann Schenk von Grafenburg
(1530-1598) published in 160090 of a central cavity in the gland,
corroborated and supplemented by Bauhin's "discovery,"
in 1605, of a black fluid in the cavity. This concept, and the
same plates, were passed on by Caspar's son, Thomas. Thomas Bartholin
(1616-1680)7, the second of six sons, was
destined to become more famous than his father. And it was Thomas'
son, Caspar the Younger, who described the greater vestibular
glands of the female perineum. An interesting history of the Bartholins,
who were well known as men of letters in addition to being famous
anatomists, is provided by Sir Humphry Davy Rolleston89.
So it is clear that by
the early 17th century, the presence of the adrenal glands in
the human abdomen had been established, although two centuries
were to pass before the medulla was unequivocally recognized.
We will return to that controversy later.
Whereas the anatomists
of this early period knew the adrenal glands were present (only
the first descriptions are mentioned above), there was great uncertainty
as to their function. This was to remain the case until the time
of Thomas Addison (1793-1860). As late as 1827, John Redman Coxe
(1773-1863) commented, "Notwithstanding, however, the
long period that has elapsed since their discovery, their use
in the animal oeconomy still remains a mystery, almost as many
offices having been attributed to them, as there have been writers
on the subject."25 Many of the early workers, probably
including Galen and certainly including Eustachius, theorized
that the adrenal glands functioned as accessory kidneys. Eustachius'
term glandulae renibus incumbentes implies this. A similar
view was hold by Julius Casserius (1545-1616), one of William
Harvey's teachers at Padua, who called them renes succenturiati.
In an interesting account given by Shumacker94,
Antonius Molinetti published a more sophisticated version of the
"renal" function of the adrenal glands in 1675. By this
time it was noticed that the adrenal glands are relatively larger,
with respect to the kidneys, in the fetus than the adult (now
known to be due to an enlargement of the cortex). It was theorized
by many that the adrenals had a more important function in fetal
life than in adulthood. Molinetti proposed that the adrenals functioned
only as diverticulae of the blood, diverting most of the arterial
blood away from the kidneys and thereby preventing the secretion
of urine in the fetus. He reasoned that this would provide some
means of avoiding urinary excretion by the fetus, since this was
considered to be disastrous. Molinetti felt that most of the arterial
blood was diverted from the kidneys, only as much reaching them
as they needed for their metabolism, the remainder being shunted
back to the heart. From here it passed through the umbilical vein
and placenta into the mother's circulation. Upon reaching the
maternal kidneys, waste material was eliminated. This explained
why women have to urinate more during pregnancy! While this theory
was virtually ignored for a century and a half, curiously it was
resurrected by Thomas Mitchell in 1813, Charles Bell in 1829,
and enthusiastically by Coxe in 1827. In supporting Molinetti's
view with persuasive reasoning (but devoid of experimental proof),
Coxe ended his case by saying if he is "...correct in
the positions thus advanced, I think all the names by which these
organs have been hitherto distinguished, should be abolished,
and that of diverticula urinae should be substituted in their
place."25
Although a renal function
for the adrenal glands was an early and long-lasting concept,
there were many other functions proposed between the times of
Eustachius and Addison. Shumacker94 gives a most thorough account
of these. Adrianus Spigelius (1578-1625), who called them capsulae
renales, thought that the adrenal glands merely served to
occupy the space between the kidneys and the diaphragm, and to
aid in supporting the stomach. This view was upheld by Nathaniel
Highmore (1613-1685) who additionally suggested that they might
serve to absorb humid exudates (sweat?) from the large vessels
nearby. Naturally such a simple theory did not satisfy many. Thomas
Bartholin replied to Spigelius by writing: "1. Nature
makes, nor does nothing in vain or inconsiderately, much less
does she appoint a noble animated Part, only to fill a space,
which she might have filled by making the kidney a little bigger.
2. These props would have been too weak by reason of their smallness.
Nor should this use belong to the Stomach alone, but to other
neighboring Parts." 7
Jean Riolan, both the
Elder (I538-1605) and his son the Younger (1577-1657), put forth
the thesis that the adrenal glands served to sustain the group
of nerves above the kidney. According to Shumacker94,
the Riolans felt that the chief need for such a support for the
nerve plexus was to prevent it from weighing too heavily upon
neighboring veins. Molinetti refuted this view by stating that
the adrenals lay above the plexus and that the plexus itself was
so firmly adherent to the vertebra that it could be freed from
it only with a scalpel. To Riolan the Younger is also attributed
the theory that the adrenals served no other purpose than to generate
perirenal fat in the fetus.
Thomas Wharton (1610-1673)
also associated the nearby nerve plexus with the function of the
adrenal glands. He referred to the adrenals as glandulae ad
plexum. As pointed out by Shumacker94,
Wharton was impressed with the large size of the plexus and with
the large nerve supply to such a small organ. He suggested that
the adrenals received some substance from the nerves (which he
assumed to be useless to the nerves); the adrenals absorbed this
substance and passed it into the veins where it had some useful
purpose. Although there is no direct evidence that Wharton supported
his hypothesis with experimentation, to him goes the credit for
being the first to associate the adrenal glands with a function
of the nervous system. Indeed his description, published in 1656,
of the adrenals taking a substance from nerves and transferring
it to veins preceded the neuroendocrine concept of the adrenal
medulla that we have only appreciated in the 20th century!
Wharton's best-known student,
Francis Glisson (1597-1677), supported his younger teacher. In
1657 he described a vessel coming from the body of the adrenal
gland that emptied into the neighboring vein. He also described
a valve in the vein which prevented return of blood to the adrenal.
Glisson thought that the adrenal glands extracted a liquid from
the nerves and secreted it into the veins. As pointed out by Shumacker94,
it would have been ridiculous to assume that the adrenals should
separate a substance from the arterial blood and then secrete
it directly back into the venous circulation. Glisson reasoned
that it must come from the nerves since they were the only other
structures communicating with the glands. Just what this liquid
was he did not know.
One of the most widely
debated of the early theories of the function of the adrenal glands
was that of Caspar Bartholin, later defended by Thomas. As mentioned
above, the Bartholins described the adrenal glands as having a
cavity filled with a brownish fluid. They held, and staunchly
defended, the belief that the adrenals absorbed atrabiliary (a
word meaning black bile, but referring to melancholy) juice from
the blood exiting the liver and spleen; from the adrenals the
juice was carried through the blood to the kidneys for excretion.
This explained, among other things, the dark color of the urine
in some disease states.
Thomas Petruccius (1648-1711)
published a book on the adrenal glands in 1675 and supported the
Bartholins' view. As discussed by Shumacker94,
Petruccius is said to have discovered valves in the capsular veins
placed in a manner contrary to those of other veins and in such
a way that they favored the passage of atrabiliary juice to the
kidneys. According to Fulton37, this rare book is also remarkable
for its inclusion of good bibliographical references to the descriptions
in question, a rather unusual thing for a 17th century medical
writer to include. However, Petruccius offered another function
for the adrenal glands. He pointed out that since "...being
a work of God Almighty, they have to contribute to the beauty
and utility of the microcosmos we call man. Like everything, they
praise God. When do they praise him? When they are viewed with
understanding eyes ......" It is no mere coincidence
that Petruccius' work was dedicated to a Cardinal.
The Bartholins were also
supported, although not enthusiastically, by Johann Vesling (Veslingius)
(1598- 1649). According to Lenard65, Vesling was an immensely popular
anatomist, based in Padua. His well- known treatise, published
in 1653, suggested that the adrenal glands "probably favour
the evacuation of the serous liquid and store the black bile which
acts as a ferment, promoting the secretion of liquid from the
blood." However, he also said of the adrenals, "What
their use is, the human mind has not yet conjectured."
There are many detractors
of the Bartholins' theory, most of them arguing on anatomical
grounds. They included Molinetti and Highmore. Another writer
of the period, Dominicus de Marchettis (1626-1688), pointed out
the obvious fact that "melancholy humor" was
especially abundant in adults, whereas the adrenal glands are
very small in adults and large in fetuses. lsbrand de Diemerbroeck
(1609-1674) wrote in favor of Vesling (and by association, the
Bartholins) but pointed out a weakness of the theory. According
to Shumacker94, de Diemerbroeck was among those
who could not demonstrate an appropriate passage from the adrenals
to the kidneys. He did conjecture that the adrenals may extract
black juice, needed by the venous blood, from the arterial blood,
thereby preventing the juice from going to parts where it is not
needed.
Theodorus Kerckringius
(1640-1693) thought that the function of the adrenal glands was
to secrete a juice to color and animate the blood and produce
a fermenta- tion in the heart. As related by Shumacker94,
this fermentation served in some manner to excite the heart. Could
Kerckringius have discovered adrenaline? Of course not, but his
theory strikes tantalizingly close to the mark.
Shumacker94 and Coxe25 mention several other writers who associated adrenal function with some essential alteration of the blood. One such group felt that its action was particularly concerned with changes in the blood of the renal circulation. Samuel Collins (1618-1710) suggested that the adrenals extracted a fermentative liquor from nerves which in turn disposed the blood to give up its waste products to the kidneys. Franciscus Sylvius (1614-1672) thought that the adrenals separated some liquor from the blood which was mixed with the blood returning from the kidneys after the secretion of urine, serving to dilute it and prevent coagulation. Hermannus Boerhaave (1668-1738) also taught that the adrenals corrected the fluidity of the blood coming from the kidneys, presumably because it was made more viscous by the extraction of urine. Antonius Deidier (?- 1746) held a similar view, maintaining that the humor from the adrenal cavity was emptied into the renal vein in order to give back to the blood a substance ("serosité") and lymph that it had lost while traversing the kidney. Daniel Tauvry (1669-1701) said that the adrenal glands strained from the blood a liquor which became oily after storage. The adrenals would "after that, throw it into the Veins; where the Blood, that was stripp'd of its Fluidity by the Secretion of the Serous Parts, recovers its former State, and its Parts that hang'd loosely together are compacted by this Liquor."
The central argument for these various theories involving a direct
effect of the adrenal glands on the blood was the observation
that the glands were hollow and contained a fluid. it is fair
to say that this description, made popular by the Bartholins,
was largely accepted as fact during the 17th and 18th centuries.
It is obvious from the foregoing descriptions that the Bartholins
alone were not responsible for this misconception. Scores of anatomists
supported the view that the adrenals were hollow. Sorkin101 suggested
that the fault should be placed on the humoral conception of medicine
and the conviction it carried in an era in which inductive reasoning
prevailed.
One of the few dissenters
from the concept of hollow adrenals during this time was Jean
Riolan (the Younger). As cited by Lenard65,
Riolan stated in 1655: "I have never seen a cavity in
them. Should they have a cavity, no pea would find a place in
it." This view was essentially ignored, even though anatomists
were having a difficult time finding the duct that they reasoned
should be draining the adrenal cavity. It was Riolan who named
the glands capsulae suprarenales, a term equivalent to
the modern one (Nomina Anatomica (Fifth edition, 1983): glandula
suprarenalis]. It is most unfortunate that Riolan is best
remembered as William Harvey's chief opponent. While he was wrong
in opposing the emerging view of the circulation, his anatomical
concept of the adrenal glands was correct. Neither of his views
found acceptance during his lifetime.
It is apparent that peas
were (and still are) a popular indication of volume. In direct
contradiction of Riolan, Gottfried Welch wrote in his thesis in
1691 that the adrenal gland contains a cavity which is able to
hold not only eight, but even twelve peas. This cavity served
as a reservoir for lymph. According to Lenard65,
Welch apparently had the courage (if that's the right word) to
taste the black liquid in the medulla and described it as "acid
and astringent." We will return later to the "discovery"
of a solid adrenal medulla.
Another category of theories of adrenal physiology involves a role in sexual function. In 1719, Antonius Maria Valsalva (1666-1723) announced the presence of an excretory duct from the adrenal communicating with the left epididymis, although Shumacker94 points out that credit for this "discovery" belongs to Marcus Aurelius Severinus (1580-1656). Valsalva also described a duct between the left adrenal and ovary. It is now apparent that he had located a left gonadal vein. Valsalva supported his view of an adrenal-gonadal relationship not only with anatomical and philosophical arguments, including Biblical references to the close association between the kidneys and genitals, but in what may have been a significant departure from the style of the day, he performed experiments to prove his point. During his career, Valsalva used several animals, including guinea pigs, birds, tortoises, and rats, but in his most famous experiment he removed one testis and the contralateral kidney (and presumably the adrenal) from a dog106. During the period of a few days before the dog died (from sepsis, no doubt), the dog did not "even fawn upon a bitch in heat." (A direct translation would read, "a bitch bitching," which has different meaning in our day.) This therefore "proved" a role for the adrenals in regulating the libido.
Although he was aware of the fact that there is no anatomical
connection between the adrenal glands and the gonads, Johanne
Friedrich Meckel (1714-1774) also associated the adrenals with
sexual function. He published in 1806 that the basis for this
relationship was "...their simultaneous, considerable
development in several orders of the mammalia...".76.
He also cited abnormalities of the adrenal glands in cases associated
with sexual abnormalities (castration, syphilis, etc.). It is
of interest to note that the direct relationship between the adrenals
and gonads, as envisioned by these early workers, had no basis
in fact, although today we do recognize important interactions
between the gonads and the adrenal cortex.
Coxe24, Shumacker94,
and Lenard65 all point out some other miscellaneous
functions that were ascribed to the adrenal glands by early investigators.
Jean Baptiste Senac (1693-1770) supposed the adrenals secreted
the fetal meconium. Jean van Helmont (1577- 1644) thought they
secreted a lithotriptic juice that prevented the formation of
renal calculi. Joseph Lieutaud (1703-1780) is said to have claimed
that the adrenals secreted an acrid liquor that prevented concretions
in the vena cava. (If it does serve this purpose, it works particularly
well in most of us!) Giovanni Morgagni (1682-1771) conjectured
that the adrenals drained lymph from the intestines during fetal
life since the cisterna chyli does not fill during this period.
In an article published in 1846, John Goodsir (1814-1867) theorized
that the adrenal glands, thymus, and thyroid have a related embryology
and may function similarly to "...elaborate the matter
which has already been absorbed by the other parts, and is now
circulating in the vessels of the more perfect individual."
One can see the germ of modern endocrinology in this concept.
Georg Heim (1803-?) suggested that the adrenal glands might serve
to divert blood away from the lungs. Evidence for this theory
included the observation that the adrenals were large in the fetus
when such a diversion took place. In 1789, Cassan made the observation
that adrenal glands were larger in Negroes than in Europeans and
offered the possibility that the adrenals played a role in pigmenting
the skin. And about this same time, Johannes Christophorus Heino
Schmidt stated that a secretion was formed in the adrenal glands
which was poured into the blood and in turn served to help the
action of the heart91. It is tempting to speculate,
although without basis, that this concept presaged the positive
inotropic effect of epinephrine on cardiac muscle.
To conclude the overview
of early concepts of adrenal function, it is instructive to note
a literary competition that was offered by the Academy of Sciences
of Bordeaux in 1716. The question the essayists were to answer
was, "What is the function of the adrenal glands?"
Although many essays were submitted, not one was considered worthy
of the prize.
As was mentioned above,
the thought of the day was strongly influenced by the concept
that the adrenal glands were hollow. Riolan was an unheard dissenting
voice. In 1797, Matthew Baille (1761-1832) referred to the soft
state, which might be almost fluid, of the center of the "renal
capsules" and commented: "This is probably what
is meant by authors when they say that they have found in the
cavity of the renal capsules a fluid like ink. Their description
may be regarded as being a little exaggerated." Although
earlier descriptions were given, the credit probably goes to Georges
Cuvier (1769-1832) for establishing the fact in 1805 that the
adrenal glands are solid structures. He appeared to recognize
that the center of the gland was morphologically distinct from
the outer portion, although the terms for the medulla and cortex
were not introduced until 1836. Meckel pointed out in 1832 that
the apparent cavity was in all probability due to postmortem softening
of the medulla, a point made in modern terms by A. David Smith
and Hans Winkler in their definitive characterization of the lysosomes
of adrenal medullary cells in 196698. Meckel differentiated the more
solid yellow substance, cortex, from the inner, softer, reddish-brown
medulla. Particularly strong evidence was presented in 1836 by
N. Nagel, who showed by means of injected preparations that the
large central vein had been mistaken for a central cavity80.
In 1839, Pierre Rayer (1793-1867) explained that the formation
of the cavity was due to extravasation of blood from the rupture
of veins. This may have been a contributing factor, but in any
event it had been established by this time that the adrenal gland
is a solid structure and that any cavity observed within was a
postmortem artifact.
In his 1836 article Nagel
used, apparently for the first time, separate terms for the two
portions of the adrenal gland. In his opening sentence, he refers
to the human adrenal gland as being composed of a cortical (Rinden)
and medullary (Mark) substance80. This terminology persists to
the present.
It was during the early
part of the 19th century that scientists were able to have a closer
look at structure. This involved the use of an improved compound
microscope, fitted with the latest in technology, achromatic lenses.
As history was to repeat itself a century and a half later when
the electron microscope was introduced, the earnest application
of the light microscope to biological studies had to await the
development of suitable preparative techniques. One of the first
to apply this new technology to the adrenal gland was Alexander
Ecker (1816-1887). Among other observations, he noted in 1846
that the adrenal medulla is remarkable for its brightness32.
However, the credit for
the first complete description of the microscopic anatomy of the
adrenal gland goes to (Rudolph) Albert von Kölliker (1817-1905).
Describing fine structure in a manner remarkable for his time,
Kölliker presented a written account in 1852 that has only
been slightly improved since60. His drawings are also remarkable.
But perhaps most impressive of all was his analysis of adrenal
function. He stated clearly: "I consider the cortical and
medullary substances as physiologically distinct." This is
a point that was apparently missed by his contemporary, Thomas
Addison. As for the different functions of the adrenal cortex
and medulla, Kölliker went on to say: "The former
may, provisionally, be placed with the so-called 'blood-vascular
glands,' and a relation to secretion assigned to it; whilst the
latter, on account of its extremely abundant supply of nerves,
must be regarded as an apparatus appertaining to the nervous system,
in which the cellular elements and the nervous plexus either exert
the same reciprocal action as they do in the grey nerve-substance,
or stand in a relation as yet wholly unascertained, towards each
other." 61 In so stating, Kölliker
was affirming Wharton's earlier (1656) idea that the adrenals
are related functionally to the nervous system. As those in the
field well appreciate, we are still trying to unravel the details
of this relationship!
The year 1856 ushered
in an era that has not yet ended, the application of histochemistry
to the adrenal medulla. These early studies provided definitive
proof that the medulla was a distinct and special part of the
adrenal gland. Gabriel Constant Colin (1825-1896) noted that the
surface of the adrenal medulla became blue when treated with sulphate
of iron21. In a more widely cited study,
(Edme Félix) Alfred Vulpian (1826-1887) showed that ferric
chloride colored the adrenal medulla green111.
This reaction was also noted by Rudolf Virchow (1821-1902) in
1857108. Perhaps more importantly, Vulpian
also noted that blood in the adrenal veins gave the same reaction,
while other organs and the blood from these organs did not. He
concluded that the material giving the reaction was related to
the function of the adrenal medulla.
The most famous histochemical
reaction associated with the adrenal medulla, the chromaffin reaction,
named for a relatively specific reaction to chromate salts, was
apparently first observed by Bertholdus Werner as a brownish deposit
after fixation in chromic acid or dichromate salts113.
Similar observations were made by Gregor Joesten in 186457
and (Friedrich Gustav) Jacob Henle (1809-1885) in 186547.
Henle was the first to give cytologic details of the color reaction.
Following these articles, there was a significant increase in
interest in the adrenal medulla. The new techniques of microscopy
were coming into general use and there was something peculiar
about the way the adrenal medulla reacted with certain metallic
solutions, particularly with chromium. However, it was not until
the end of the century that the term "chromaffin" was
introduced to describe this reaction. The term "chromaffin
reaction" was applied to cells that reacted in this manner,
the adrenal medulla being the prime example. The "chromaffin
cell" and apparently the name of the reaction as well, were
so named by Alfred Kohn (1867-1959) by 190259.
(A brief story of how Kohn survived in the concentration camp
at Theresienstadt is provided by Blaschko10.)
These and related studies
in the last half of the 19th century clearly established that
the adrenal medulla is histologically and functionally separate
from the cortex. What is now known as "chromaffin tissue"
was coming into its own. Interestingly, there have been several
other terms introduced with which to label this tissue. These
proposed terms include "chromophile,"102,
"pheochrome,"86 and "fuscogenic"8.
While these and other terms have merit, the term "chromaffin"
has stuck and is used universally to describe cells of the adrenal
medulla and related cells that exhibit the chromaffin reaction.
While this discussion
of the histochemistry of the adrenal medulla has only covered
the earliest studies (for more current and much more comprehensive
information, see reviews by Rex Coupland24
and David Hopwood54), a particularly interesting
article was published in 1918. William Cramer (1873-1930) showed,
but not for the first time, that osmic acid had a specific effect
on the adrenal medulla. He noted that osmic acid vapor demonstrated
"adrenaline granules" as "giving the
appearances of fine coal dust scattered over the medulla"26.
He was probably the first to give clear evidence that an increase
in secretary activity is a result of the passage of these osmiophilic
granules into the blood vessels. It has been shown, and continues
to be shown, that this observation describes the essential event
in the secretion of catecholamines from the adrenal medulla. Only
the tools used to show it have changed.
During the middle of the 19th century, embryologists began to examine the development of the adrenal glands. In 1831, Friedrich Arnold (1803-1890) stated that the adrenals developed from the Wolffian (mesonephric) bodies by means of a fissure, and that there was a structural resemblance between the two2. In a rather remarkable series of articles, beginning in 1847, Robert Remak (1815-1865) indicated that the adrenal medulla originated in association with the sympathetic ganglia88. During his brief and extremely productive career, Henry Gray (1827-1861) also investigated the embryology of the adrenal glands. In a paper on "the ductless glands of the chick" published in 1852, he reported that the adrenals arose from "two grayish white masses, which lie one on each side of the aorta, between this tube and the upper and inner extremity of the Wolffian bodies, and are perfectly separate"42.
Kölliker showed that
the adrenal cortex is formed first and is subsequently invaded
by sympathetic neural elements. Although there were several important
studies on comparative embryo- logy of the adrenal glands (see
Rolleston89, for review), these early observations
were scarcely improved upon until the already-classic studies
of chimeras by Nicole Marthe Le Douarin (1930- ) and her colleagues
in the 1970s (for example, see64). For a review of more recent
work, see18.
The stage was now set
for the breakthrough that would give us an understanding of what
the adrenal medulla actually does. By the end of the 19th century
th basic anatomy, histology, histochemistry, and embryology of
the adrenal medulla were understood. It was appreciated both on
histological and embryological grounds that the adrenal medulla
was somehow associated with the nervous system, specifically the
sympathetic nervous system. The classic story of this breakthrough
puts George Oliver and Edward Albert Schäfer at center stage.
Perhaps this is where they belong but, as we shall see, there
are some challengers. As for Oliver (1841-1915) and Schäfer
(1850-1935), Sir Henry Dale (1875-1968) gives a charming account
of the circumstances surround- ing their discovery27.
Oliver, a physician of Harrogate, employed his winter leisure
in experiments on his family, using equipment he had designed
for clinical testing. In one such experiment he was applying an
instrument for measuring the thickness of the radial artery, and
having given his young son (whom Dale suggested should receive
a special memorial!) an injection of an extract of the adrenal
gland prepared from material supplied by the local butcher, Oliver
thought that he detected a contraction (others suggest it was
an expansion) of the radial artery. When Oliver went to London
to tell Professor Schäfer what he thought he had observed,
he found Schäfer engaged in an experiment in which the blood
pressure of a dog was being recorded. Schäfer was, not unnaturally,
incredulous about Oliver's claim and was very impatient at the
interruption. But Oliver was in no hurry, and urged only that
a dose of his adrenal extract, which he produced from his pocket,
should be injected into the dog when Schäfer's own experiment
was over. And so, just to convince Oliver that it was all nonsense,
Schäfer injected the extract into a vein, and then stood
in amazement as they watched the mercury climbing in the arterial
manometer until the recording float almost lifted out of the distal
limb. Their publication in 189484 resulting from their subsequent
experiments is heralded as the first demonstration of a hormonal
effect. Many historians regard this study of the adrenal medulla
as a milestone in endocrinology.
As pointed out by Medvei77,
another scientific team made the same discovery independently
of Oliver and Schäfer. Napoleon Cybulski (1854-?) and Ladislaus
Szymonowicz performed experiments quite similar to Oliver and
Schäfer's in Cracow in December 1894, just one year after
the first experiment in London103.
However, the first indirect
demonstration of the role of the adrenal medulla as an endocrine
organ was in 1892. Joh. Carl Jacobj (1867-?) showed in 1892 that
electrical stimulation of the greater splanchnic nerve in the
dog decreased the amplitude of contraction of the animal's intestines56.
Whereas this article appears not to have been appreciated at the
time, hindsight shows us that this was actually a more sophisticated
demonstration of adrenal medullary function than the classic study
of Oliver and Schäfer.
Even before Jacobj's publication,
there was another clue to the function of the adrenal medulla.
In 1886, Felix Fränkel reported the first case of a tumor
of the adrenal medulla35, a type of tumor that has become
known as pheochromocytoma. The patient in this case was an 18
year-old girl who had died suddenly of collapse. Her clinical
history and autopsy findings pointed to a severe hypertensive
crisis. This, combined with the discovery of an adrenal medullary
tumor, presented what appears to be the first evidence, seer-,
only in retrospect, of the relationship between the adrenal medulla
and blood pressure.
There are some accounts
of the history of pathologies of the adrenal glands (for example,
Sorkin101 and Rolleston89)
and pheochromocytoma in particular (Manger and Gifford74,
Sherwin93, Medvei77,
and Welbourn112). Just a few highlights of this
history will be given here. After Fränkel's report, Paul
Manasse (1866-?) reported another patient with pheochromocytoma
in 189371. In 1896, Manasse was the first
to demonstrate that chromium salts reacted with pheochromocytomas73.
In the ensuing years, there was a heated controversy over the
nature of this reaction, both with tumors and normal adrenal medullary
tissue. Heinrich Poll (?-1939) objected to the word "chromaffin"
to describe this reaction because of the uncertain nature of the
dark color produced. (Blaschko10 gives an interesting account
of Poll's life and death.) Poll introduced the term "pheochrome,"
and in 1912, Ludwig Pick (1868-1935) incorporated this into the
neoplastic counterpart of the adrenal medulla. The term "pheochromocytoma"
has been used ever since.
In the early part of this century the clinical picture of pheochromocytoma was emerging. It wasn't until 1926, however, that surgical intervention was attempted. The first case of a successful removal of a pheochromocytoma was performed by César Roux (1857-1934) (cited by Manger and Gifford74 and Welbourn112; this report was not published in the traditional sense, but is apparently only recorded in a thesis at the University of Lausanne). About 7 months later, Charles Horace Mayo (1865-1939) also removed a pheochromocytoma75. The patient in this historic case was Mother Joachim. A fascinating account of Mother Joachim, and her recovery after this surgery, is provided by van Heerden107.
Maurice Charles Pincoffs
(1886-1960) is credited with first recognizing a pheochromocytoma
preoperatively85. In the same year, 1929, Coleman
Berley Rabin (1900- ) noted that a pheochromocytoma contained
a pressor agent in excess of that occurring in the normal adrenal
medulla and suggested that this might account for the clinical
manifestations87. Although several significant
features of pheochromocytoma have been published in the meantime
(see, for example74,112 for details), the essential characteristics
of the principal adrenal medullary tumor had been recognized by
this time.
Alongside these studies of the clinical picture of adrenal medullary
pathology, physiologists were carrying out the classic studies
on the function of the adrenal medulla. Independently, George
P. Dreyer (1866-1931) in 189830
and M. Lewandowsky
in 189967 noted the correspondence between
the effects of stimulation of postganglionic sympathetic nerves
and the effects of administered epinephrine. This was confirmed
by John N. Langley (1852-1925)63, whose student, Thomas Renton
Elliot (1877-1961), suggested in 1904 that the sympathetic nerves
produced their effects by liberating epinephrine33.
This suggestion is considered by many to be the beginning of our
concept of neurochemical transmission.
Although there were many
other great scientists pursuing the problem of adrenal medullary
function in the first part of this century, the name that stands
out is Walter Bradford Cannon (1871-1945). It is largely through
the efforts of Cannon and a host of collaborators (see the career-end
review by Cannon13) that we have a basic understanding
of how the adrenal medulla contributes to the body's efforts to
maintain homeostasis during stress. In fact, the still-used term
"stress syndrome" was coined by one of Cannon's students,
Hans Selye (1907-1982). However, Cannon and his contemporaries
encountered phenomena that could not be explained by just the
elaboration of epinephrine from the adrenal medulla. The discovery
of norepinephrine was not made until after Cannon's death. Furthermore,
as pointed out by Davenport28, Cannon used a commercial preparation
of natural epinephrine that contained norepinephrine.
Also during the early
part of this century, there was a battle raging in the chemical
laboratories. Davenport28 tells the intricate tale of the
struggle for priority as the discoverer of epinephrine, a struggle
particularly between John Jacob Abel (1857-1938) and Jokichi Takamine
(1854-1922), but also involving Thomas Bell Aldrich (1861-?).
The story is rife with intrigue and discovery, accusations and
counter-accusations. A suggestion of how complicated the picture
gets is offered by the fact that the Merck Index 178 lists 38
different terms for what is generally called epinephrine. This
includes such interesting names as Takamine and Supracapsulin.
Incidentally, the term used in the present account is epinephrine,
as listed in the United States Pharmacopeia. The widely used term,
adrenaline, is listed in the British Pharmacopoeia.
Ernst Joseph Friedmann
(1877-1956) published the chemical formula of epinephrine in 190636.
An account of Friedmann's interesting life is given by Mitchell79.
One of the first schemes of the biosynthetic pathway for epinephrine
was proposed by Walter L. Halle, also in 190645.
It is interesting to note that Halle's scheme differs very little
from that proposed by Hermann Karl Felix Blaschko (1900- ) in
19399. Blaschko's pathway was proven
to be correct in 1957 by McChesney Goodall (1916- ) and Norman
Kirshner (1923- )41. An account of how the enzymes
of the pathway (tyrosine hydroxylase, dopa decarboxylase, dopamine
ß-hydroxylase, and phenylethanolamine N-methyl transferase)
were discovered is given by Hingerty and O'Boyle52.
The discovery of norepinephrine
(noradrenaline), the biosynthetic precursor of epinephrine and
a bioactive compound in its own right, as a secretary product
of the adrenal medulla was a particularly important break-through.
To be sure, there were many scientists involved in the elucidation
of the role of norepinephrine in the sympathetic nervous system,
but the most prominent of these was Ulf Svante von Euler (1905-1983).
His contributions to the field of pharmacology were recognized
with the Nobel Prize in 1970. As an example, in 1949 von Euler
was among the first to demonstrate that norepinephrine is a constituent
of the adrenal medulla110. This report was followed in
5 weeks by an independent report by Marcel Goldenberg and colleagues
who reached the same conclusion39. However, the first report of
norepinephrine in the adrenal medulla was made some months earlier
by Peter Holtz (1902-1970)53. It is of interest to note that
these reports came just a few years after Cannon's death. Cannon
would have loved to have heard this news.
The next big breakthrough
came just a few years later. Under the light microscope the cytoplasm
of adrenal chromaffin cells appears granular due to the presence
of numerous organelles, often referred to as chromaffin granules.
Chromaffin granules were apparently first noticed by Manasse in
189472. Numerous microscopists made
note of the granular cytoplasm in subsequent years, but the definitive
study of the adrenal medulla at the fight microscopic level was
published in 1941 by Henry Stanley Bennett (1910-1997)8.
With remarkable resolution, he demonstrated chromaffin granules
that appeared to be ripe for the plucking. And plucked they were.
In 1953, two independent laboratories reported the isolation of
chromaffin granules from the bovine adrenal medulla. Blaschko
and Arnold D. Welch (1908- )12 and Nils-Åke Hillarp (1916-1965)
and his colleagues50 submitted papers exactly one
month apart. Both groups showed that centrifugation of homogenates
of the adrenal medulla led to the sedimentation of the major part
of the hormones of this organ, the catecholamines. This was the
first time a secretary organelle was isolated. Subsequently, the
concept of the chromaffin granule as a distinct hormone-containing
component of the cytoplasm also paved the way for the discovery
of hormone- and neurotransmitter- containing organelles in other
organs. As pointed out by Winkler and Carmichael118,
it is not an overstatement to say that the first-discovered secretary
organelle remains the best characterized and has often served
as a "model" organelle. (For further discussion, including
600 references to more recent work, see118).
Shortly after the isolation
of chromaffin granules using biochemical techniques, the electron
microscope was used in this area. The first relevant application
of the electron microscope was by Hillarp and his colleagues49
who examined smears of crushed medullary cells in 1954. The first
ultrastructural study of the adrenal medulla was published by
Jeffrey Darcy Lever(1923- ) in 195566. In 1956, Fritiof Stig Sjöstrand
(1912- ) and Rudolph Wetzstein (1916- )97
were the first to use the term "chromaffin granule"
to describe the catecholamine- containing organelle of the adrenal
chromaffin cell. Although this organelle has been clearly shown
to be a membrane-bound cavity, a structure properly referred to
as a vesicle, Sjöstrand still feels the term granule is preferable
(personal communication). The terms "chromaffin granule"
and "chromaffin vesicle" are used interchangeably in
the literature.
Another important contributor
to our understanding of the morphology of the adrenal chromaffin
cell has been Rex Ernest Coupland (1924- ). His 1965 book on the
natural history of the chromaffin cell24 has remained a classic although,
alas, it has been out of print for several years. Since these
first electron microscopic studies, the ultrastructure of the
adrenal medulla has been described in more than 50 animals (for
references, see16). The adrenal medulla has also
been studied with both the high-voltage (1000 KV)17
and the scanning electron microscopes19.
Up until the work of the
mid-1950's, the centuries of work on the adrenal glands and the
adrenal medulla had been directed toward the anatomy and physiology
of the entities themselves. From the 50's onward, the emphasis
shifted from a study of adrenal medullary function per se to an
outlook where the adrenal chromaffin cell was regarded as a model
neuron in general, a model sympathetic neuron in particular. This
view is held to the present day, and is sure to direct research
on the adrenal medulla in the future.
There are several examples
of how the adrenal medulla has served as a neurobiological model.
One is the phenomenon of the uptake of compounds into cytoplasmic
vesicles, whether they be chromaffin vesicles, synaptic vesicles,
or another organelle such as the lysosome. It was noticed in 1962
that chromaffin granules can take up epinephrine from a dilute
solution against a concentration gradient provided that the medium
contains adenosine triphosphate (ATP) and magnesium ions14,58,
that is, the uptake of epinephrine into chromaffin granules is
an active process. It was shown a short time later by Peter Banks(1937-
) that a magnesium-activated ATPase is associated with the chromaffin
vesicle, suggesting the uptake of catecholamines is linked to
the hydrolysis of ATP by this enzyme5. A decade was to pass before
this link was elucidated. George Karoly Radda (1936- ), David
Lars Njus (1948- ) and their colleagues82,
and independently, Robert Gahagen Johnson (1952- ) and Antonio
Scarpa (1942- ) demonstrated this to be a coupled transport system.
More specifically, the ATPase actively drives protons into the
chromaffin vesicle and the proton gradient (creating pH and electrical
gradients across the vesicle membrane) in turn drives catecholamine
uptake (see Njus and Radda82 for details). This was the first
demonstration of a chemiosmotic mechanism operating in any mammalian
organelle other than the mitochondrion. The usefulness of the
adrenal medulla in the discovery of this important mechanism (it
has since been shown to operate in several other systems) led
Njus et al.81 to make the following whimsical
proposal about "the legacy of the adrenal medulla":
"Often the unrelenting pressure of evolution creates features
so exaggerated they seem intended more for the edification of
biologists than for the survival of the organism. The adrenal
medulla is a good example." While this may not be literally
true, it certainly makes good press!
Another example of the
value of the adrenal medulla in neurobiology has been studies
on the composition of the chromaffin vesicle since this serves
as a model neurotransmitter-containing vesicle. The pioneering
work of Hillarp in 1959 demonstrated that proteins and nucleotides
are contained within the chromaffin vesicle48,51.
In 1966, Karen Blaauw Helle (1934- )  published
the first attempt to characterize the soluble proteins of chromaffin
vesicles in some detail, and also described the first preparation
of antibodies against the soluble protein of chromaffin vesicles.
These antibodies were used for the first demonstration of exocytosis
by Banks and Helle in 19656. Within a year, Blaschko's group
had given the name "chromogranin" to a protein within
chromaffin vesicles and had shown that it was secreted from the
adrenal medulla11, although the name "chromogranin"
was first suggested by Helle (personal
communication). Anthony
David Smith (1938- ) and Hans Christoph Winkler (1939- ), working
in Blaschko's laboratory, and, independently, Kirshner, purified
chromogranin A (as it is now called)99,100. Impressive progress has been
made in the last 20 years in our understanding of the composition
of the chromaffin vesicle, but important questions remain about
the structure of the vesicle and the "cocktail" that
it secretes (see Winkler et al.118 for review).
published
the first attempt to characterize the soluble proteins of chromaffin
vesicles in some detail, and also described the first preparation
of antibodies against the soluble protein of chromaffin vesicles.
These antibodies were used for the first demonstration of exocytosis
by Banks and Helle in 19656. Within a year, Blaschko's group
had given the name "chromogranin" to a protein within
chromaffin vesicles and had shown that it was secreted from the
adrenal medulla11, although the name "chromogranin"
was first suggested by Helle (personal
communication). Anthony
David Smith (1938- ) and Hans Christoph Winkler (1939- ), working
in Blaschko's laboratory, and, independently, Kirshner, purified
chromogranin A (as it is now called)99,100. Impressive progress has been
made in the last 20 years in our understanding of the composition
of the chromaffin vesicle, but important questions remain about
the structure of the vesicle and the "cocktail" that
it secretes (see Winkler et al.118 for review).
Studies on the proteins
of the chromaffin vesicle have more recently been extended to
enkephalins. Since the discovery of enkephalins in the pig brain
in 1975, there has been an intense interest in these and related
peptides. Enkephalin-like reactivity was first demonstrated in
the adrenal medulla by Tomas G.M. Hökfelt (1940- ) and his
colleagues92. Within a year, studies in the
laboratories of Osvaldo Humberto Viveros (1937-), Sidney Udenfriend
(1918- ), and Erminio Costa (1924- ) demonstrated that enkephalins
and/or opioid peptides were localized in the adrenal medulla,
specifically within chromaffin vesicles23,68,109.
Work in these and other laboratories showed that the adrenal medulla
is one of the richest sources of enkephalins. Therefore the adrenal
medulla has been used extensively in studies on opioid peptides.
These include the virtually simultaneous publication of the sequence
of the preproenkephalin molecule by three independent groups (Noda
et al.83, Gubler et al.44,
Comb et al.22). All three groups used the adrenal
medulla as the source for material. The adrenal medulla continues
to be used extensively for studies on peptide synthesis and processing.
The utilization of adrenal
chromaffin cells in neurobiology has been increased by the use
of isolated cells. The first successful move in this direction
was the discovery by Arthur Steven Tischler (1946- ) and Lloyd
Asher Greene (1944- ) of a culturable form of the rat pheochromocytoma
cell. This cell line (referred to as PCI2 cells) has proven to
be very useful and popular (see43 for references). A recent technique
that has helped make the adrenal medulla even more valuable for
studying peptides, receptors, and many other aspects of neurobiology
was the development of methods for isolating and culturing adrenal
chromaffin cells. Among the many people who pioneered these techniques
are Jack Carlton Brooks (1941- ), Allan Stanford Schneider (1940-
), Robert Louis Perlman (1938- ) and Bruce Grayson Livett (1943-
) (see Livett69 for details and references). Jack
Brooks is known to his friends as "Smilin' Jack" 
and he's always a favorite
of the ladies! Adrenal chromaffin cells are now being cultured
around the world for use in an extensive variety of studies.
Just one more example
of current studies on chromaffin cells. In the 1960's, William
Wilton Douglas (1922- ) and his colleagues determined that calcium
ions are the only ion required for the secretion of catecholamines
from the adrenal medulla, although the dependence on calcium had
been demonstrated earlier (by Houssay and Molinelli55).
Douglas compared this to excitation-contraction coupling in muscle
and coined the term "stimulus-secretion coupling"
in 196829. The precise role of calcium
in this process was not understood then, and it is not understood
now (for review, see Baker and Knight4). Measurements of calcium levels
in chromaffin cells, using numerous techniques, have been made
by many investigators. Many intracellular proteins and other compounds
have been implicated; but we still do not know how a rise in cellular
calcium levels leads to the release of the chromaffin vesicle
contents at the cell surface (a process known as exocytosis).
When the answer comes, it will probably be with studies using
isolated chromaffin cells.
Two unrelated observations
have led to the answer of an old question about the adrenal gland.
The first of these observations was made by Richard Jay Wurtman
(1936- ) and Julius Axelrod (1912- ) (who won the Nobel Prize
along with von Euler in 1970). In 1965, they found that adrenal
cortical steroids enhance the conversion of norepinephrine to
epinephrine119. Within the past year, it has
also been shown that cortical steroids have a direct effect on
the peptides (LaGamma and Adler62), including enkephalins (Yoburn
et al.120), chromogranins and other proteins
(Sietzen et al.95).
The second observation,
made independently by Klaus Unsicker (1942- ), Jose-Maria Trifaró
(1936- ),  their colleagues and others, was
that when adrenal chromaffin cells are grown in culture they extend
neurite-like processes104,105. This growth is inhibited by
steroids in the culture medium. Work in several laboratories has
shown that the phenotypic expression of adrenal chromaffin cells
is influenced by glucocorticoids (for example, Anderson and Axel1).
their colleagues and others, was
that when adrenal chromaffin cells are grown in culture they extend
neurite-like processes104,105. This growth is inhibited by
steroids in the culture medium. Work in several laboratories has
shown that the phenotypic expression of adrenal chromaffin cells
is influenced by glucocorticoids (for example, Anderson and Axel1).
With these and related
studies in a number of laboratories, a longstanding question has
satisfactorily been answered (although additional answers are
certain to surface in the future). The question had been: Why
are the adrenal glands composed of two organs, the medulla and
cortex, that are distinct in their morphology, physiology, embryology,
etc.? The two current answers are: first, the cortical hormones
influence the activity of the enzyme that converts norepinephrine
to epinephrine (phenylethanolamine N-methyltransferase),
creating the situation whereby the medulla secretes the generally
more active catecholamine into the bloodstream. In a related fashion,
the cortex appears to influence the peptide and protein content
of the adrenal medulla. Second, cortical hormones also influence
the shape of the adrenal chromaffin cells, somehow preventing
them from extending processes, as do other postganglionic sympathetic
neurons.
For many of us working with the adrenal medulla, the adrenal chromaffin cell is considered as a model neuron and, from the hints offered above, with good reason (for a more general review, see20). More recently, the adrenal chromaffin cell has been considered as a substitute neuron in the human brain! In 1982, parts of the adrenal medulla were transplanted autologously into the brain of a patient with severe Parkinson's disease (Backlund et al.3). It has been shown more recently that, in selected patients, this may be the first effective therapy for this common disease (Madrazo et al.70).
1552 Eustachius completes plates illustrating adrenal glands.
1611 Bartholin describes hollow adrenal glands.
1655 Riolan asserts adrenal glands not hollow.
1656 Wharton associates adrenal glands with nerve plexus.
1805 Cuvier establishes that adrenal glands are solid.
1831 Arnold studies embryology of adrenal glands.
1836 Nagel names adrenal medulla.
1852 Kölliker describes microscopic anatomy of adrenal medulla.
1856 Histochemical studies of adrenal medulla begun.
1886 Fränkel describes tumor of adrenal medulla.
1892 Jacobj shows adrenal medulla is an endocrine organ.
1894 Oliver and Schäfer describe pressor effect of adrenal
medulla.
1896 Cybulski and Szymonowicz also describe pressor effect.
1901 Epinephrine discovered independently by Abel and Takamine.
1902 Kohn names chromaffin cells.
1906 Friedmann publishes chemical formula of epinephrine.
1926 First surgical removal of pheochromocytoma (Roux and Mayo).
1939 Blaschko determines biosynthetic pathway of epinephrine.
1948 Holtz detects norepinephrine in adrenal gland.
1953 Chromaffin vesicles isolated.
1955 Lever describes ultrastructure of adrenal medulla.
1968 Douglas coins terms "stimulus-secretion coupling."
1979 Enkephalin measured in adrenal medulla.
1982 Adrenal medulla transplanted into human brain.
Above is an outline of some of the developments that relate to the history of the adrenal medulla. It is of interest to note that the adrenal glands have been recognized for just over four hundred years. For almost half of that period, the glands were thought to be hollow! Since this is an article on history, it is not pertinent to discuss all of the recent and current studies on the adrenal medulla. Although many have been mentioned above, there are other areas of current activity. These include investigations on: mechanisms of exocytosis; the structure and function of catecholamine-synthesizing enzymes; the variety of receptors on chromaffin cells; the regulation of the adrenal medulla by the central nervous system; the role of the adrenal medulla in the metabolism of the organism; and the diagnosis and clinical treatment of tumors of the adrenal medulla (mainly pheochromocytomas) (see15 for references).
There are many more of
our contemporaries who have made and are making important contributions
to the history of the adrenal medulla. It is impossible to list
them all here. By way of illustration, it is documented that over
2,500 scientists authored or co-authored articles relating to
the adrenal medulla in the years 1983-1985 alone!15
As large and as talented as this field is, one worker stands out
for having made consistently important and accurate discoveries,
in addition to calling the attention of the scientific community
to the value of the adrenal chromaffin cell as a neurobiological
model. This acknowledged leader in the field is Hans Winkler.
 Winkler has been involved in most
of the significant work relating to the adrenal medulla for the
last 30 years. Furthermore, he has shown a knack for analyzing
the literature and trends, often pointing the way for others.
For example, commentaries he has published in Neuroscience on
the composition of the chromaffin vesicle in 1976114
and the biogenesis of the chromaffin vesicle in 1977115
are already regarded as classics. His 1976 article has been cited
more frequently than any other article in the journal Neuroscience116.
In his 1977 article in this journal, he made several predictions
regarding the origin and fate of chromaffin vesicles; essentially
they have all been shown to be true. Winkler and his colleagues
continue to be major contributors to our understanding of the
composition and life cycle of the chromaffin vesicle, as well
as the biological significance of the adrenal chromaffin cell117.
There can be no doubt that Hans Winkler has done more than anyone
in promoting interest in the adrenal medulla! And when he drills
you with his famous, questioning stare
Winkler has been involved in most
of the significant work relating to the adrenal medulla for the
last 30 years. Furthermore, he has shown a knack for analyzing
the literature and trends, often pointing the way for others.
For example, commentaries he has published in Neuroscience on
the composition of the chromaffin vesicle in 1976114
and the biogenesis of the chromaffin vesicle in 1977115
are already regarded as classics. His 1976 article has been cited
more frequently than any other article in the journal Neuroscience116.
In his 1977 article in this journal, he made several predictions
regarding the origin and fate of chromaffin vesicles; essentially
they have all been shown to be true. Winkler and his colleagues
continue to be major contributors to our understanding of the
composition and life cycle of the chromaffin vesicle, as well
as the biological significance of the adrenal chromaffin cell117.
There can be no doubt that Hans Winkler has done more than anyone
in promoting interest in the adrenal medulla! And when he drills
you with his famous, questioning stare you
won't forget it...
you
won't forget it...
It is tempting, but impossible, to tell the future history of the adrenal medulla. While some have expressed the feeling that we are nearing the end of important work with this system, that the relatively simple adrenal chromaffin cell will soon yield all of its secrets, I disagree. As with all good science, as the important questions of today (and only a few general ones have been alluded to in this article) are answered, these discoveries will themselves pose new questions. The adrenal medulla will continue to be a useful model for study. The history of the adrenal medulla will continue to be an important part of the history of neuroscience.
The author thanks Miss Ruth Mann who, as History of Medicine Librarian
at Mayo Clinic, urged the author to pursue this project. I also
thank Mrs. Nancy Rucker- Johnson, History of Medicine Librarian,
who has tracked down numerous articles and bits of information.
Also, Mr. Robert Spinner, one of my students, has been invaluable
in translating from Latin and Greek. Finally, I thank my colleagues
who have supplied information and/or have reviewed the manuscript
in its formative stages, including Stephen Briimijoin, Jack Brooks,
Frank Mann, David Njus, Steve Sommer, Susan Stoddard, Jon van
Heerden, Richard Welbourn, and Hans Winkler.
1. Anderson DJ, Axel R. A bipotential neuroendocrine precursor
whose choice cf cell fate is determined by NGF and glucocorticoids.
Cell 1986; 47. 1079-90.
2. Arnold F. Der Kopftheil des vegetativen Nervensystems. Salz med Zeit 1831; 301.
3. Backlund E-0, Granberg P-0, Hamberger B, Sedvall G, Sieger A, Olson L. Transplantation of adrenal medullary tissue to striatum in parkinsonism: First clinical trials. J Neurosurg 1985; 62. 169-73.
4. Baker P, Knight D. Calcium controls exocytosis in bovine adrenal medullary cells. Trends Neurosci 1984; 7: 120-26.
5. Banks P. The adenosine-triphosphatase activity of adrenal chromaffin granules. Biochem J 1965; 95: 490-96.
6. Banks P, Helle K. The release of protein from stimulated adrenal medulla. Biochem J 1965; 97: 4OC-41C.
7. Bartholin T. Bartholin Anatomy, Made from the Precepts of his Father, and from the Observations of all Modern Anatomists, Together with his Own. Nich. Culpeper and Abdiah Cole, publishers, London, 1668.
8. Bennett HS. Cytological manifestations of secretion in the adrenal medulla of the cat. Am J Anat 1941; 69: 333-81.
9. Blaschko H. The specific
action of 1-dopa decarboxylase. J Physiol (Lond) 1939; 96: 5OP-51P.
10. Blaschko H. Reminiscences, Catecholamines: Basic and Peripheral
Mechanisms. New York: Alan R. Liss, 1984; pp. 3-4.
11. Blaschko H, Comline RS, Schneider FH, Silver M, Smith AD.
Secretion of a chromaffin granule protein, chromogranin, from
the adrenal gland after splanchnic nerve stimulation. Nature 1967;
215: 58-59.
12. Blaschko H, Welch AD. Localization of adrenaline in cytoplasmic
particles of the bovine adrenal medulla. Naunyn-Schmiedebergs
Arch Exp Pathol Pharmacol 1953; 219: 17-22.
13. Cannon WB. The adrenal medulla. Bull NY Acad Med 1940; 16:
3-13.
14. Carlsson A, Hillarp NA, Waldeck B. A Mg2+-ATP dependent storage
mechanism in the amine granule of the adrenal medulla. Med Exp
1962; 6: 47-53.
15. Carmichael SW. The Adrenal Medulla, Volume 4. New York. Cambridge
University Press, 1986.
16. Carmichael SW. Morphology and innervation of the adrenal medulla.
In: Rosenheck K, Lelkes PI, eds, Stimulus-Secretion Coupling in
Chromaffin Cells. Boca Raton: CRC Press, 1987.
17. Carmichael SW, Smith DJ. High-voltage electron microscopy
of the cat adrenal medulla. Experientia 1974; 30: 1459-60.
18. Carmichael SW, Spagnoli DB, Frederickson RG, Krause WJ, Culberson
JL. The opossum adrenal medulla. 1. Postnatal development and
normal anatomy. Am J Anat 1987; 179: 211-19.
19. Carmichael SW, Ulrich RG. Scanning electron microscopy of
the mammalian adrenal medulla. Mikroskopie 1983; 40: 53-64.
20. Carmichael SW, Winkler H. The adrenal chromaffin cell. Scientific
American 1985; 253: 40-9.
21. Colin G. Traité de physiologic comparée. Paris,
1856.
22. Comb M, Seeburg PH, Adelman J, Eiden L, Herbert E. Primary structure of the human Met- and Leu-enkephalin precursor and its mRNA. Nature 1982; 296; 663-66.
23.Costa E, Di Giulio
A, Fratta W, Hong J, Yang H-YT. Interactions of enkephalinergic
neurons in CNS and periphery. In Usdin E, Kopin IJ, Barchas J,
eds, Catecholamines- Basic and Clinical Frontiers. New York: Pergamon
Press, 1979; pp. 1020-25.
24. Coupland RE. The Natural History of the Chromaffin Cell. London.
Longmans, 1965.
25.Coxe JR. On the functions of the capsulae renales. Am j Med Sci 1827; 1: 4049.
26. Cramer W. Further observations on the thyroid-adrenal apparatus. A histochemical method for the demonstration of adrenalin granules in the suprarenal gland. J Physiol 1918; 52: 7-10.
27. Dale H. Accident and opportunism in medical research. Brit Med J 1948; 2: 451-55.
28. Davenport HW. Epinephrin(e).
Physiologist 1982; 25; 76-82.
29. Douglas WW. Stimulus-secretion coupling. The concept and clues
from chromaffin and other cells. Brit J Pharmacol 1968; 34: 451-74.
30. Dreyer GP. On secretary nerves t6 the -suprarenal capsules.
Am J Physiol 1898; 2: 203-19.
31.DuLaurens (Laurentius) A. Historia anatomica humani corporis
et singularum eius partium multis controversies observationibus
illustrate. Paris: M. Orry, 1640 (from Lenard).
32.Ecker A. Der feinere Bau der Nebenniere beim Menschen und den
vier Wirbelthierclassen. Braunschweig, 1846.
33. Elliot TR. On the action of adrenaline. J Physiol (Lond) 1904;
31: XX-XXI.
34.Eustachi Bartolomeo. Tabulae anatomicae clarissimi viri bartholomaei
Eustachii quas, 1563. (Republished in Rome in 1714 by Jo. Maria
Lancisius, F. Gonzagae, publisher.)
35. Frankel F. Ein Fall von doppelseitigem vdllig latent verlaufenen
Nebenneirentumor und gleichzeitiger Nephritis mit Vergnderungen
am Circulationapparat und Retinitis. Virchows Arch Pathol Anat
Physiol 1886; 103: 244-63.
36.Friedmann E. Die Konstitution des Adrenalins. Beitr z chem
Phys und Path 1906; 8: 95-120.
37. Fulton JP. Petruccius on the adrenals. J lest Med 1956; 1
1: 224-25.
38. Galen. Siebén Bilcher Anatomie des Galen (translated
into German by M. Simon). Leipzig: J.C. Hindrichs'schue Buchhandlung,
1906.
39. Goldenberg M, Faber M, Alston EJ, Chargaff EC. Evidence for
the occurrence of nor-epinephrine in the adrenal medulla. Science
1949; 109: 534-35.
40. Goldzieher MA. The Adrenal Glands in Health and Disease. Philadelphia:
F.A. Davis Co., 1944.
41. Goodall M, Kirshner N. Biosynthesis of adrenaline and noradrenaline
in vitro.J Biol Chem 1957; 226: 213-21.
42. Gray H. On the development of the ductless glands of the chick.
Phil Trans Roy Soc Lond 1852; 142. 295-310.
43. Greene LA, Tischler AS. PC 12 pheochrormocytoma cultures in
neurobiological research. Adv in Cell Neurobiol 1982; 3: 373-414.
44. Gubler U, Seeburg P, Hoffman BJ, Gage LP, Udenfriend S. Molecular
cloning establishes proenkephalin as precursor of enkephalin-containing
peptides. Nature 1982; 295: 206-08.
45. Halle WL. Über die Bildung des Adrenalins im Organismus.
Beitr Chem Physiol Pathol 1906; 8: 276-80.
46. Helle KB. Some chemical and physical properties of the soluble
protein fraction of bovine adrenal chromaffin granules. Mol Pharmacol
1966; 2: 298-310.
47. Henle J. Ueber das Gewebe der Nebenniere und der Hypophyse.
Z rat Med 1865; 24: 143-52.
48. Hillarp N-Å. Further observations on the state of catechol-
amines stored in the adrenal medullary granules. Acta Physiol
Scand 1959; 47: 271-79.
49. Hillarp N-Å, Hökfelt B, Nilson B. The cytology
of the adrenal medullary cell with special reference to the storage
and the secretion of the sympathomimetic amines. Acta Anat 1954;
21: 155-67.
50. Hillarp N-Å,
Lagerstedt S, Nilson B. The isolation of a granular fraction from
the suprarenal medulla, containing the sympathomimetic catecholamines.
Acta Physiol Scand 1953; 29: 251-63.
51. Hillarp N-Å, Thieme G. Nucleotides in the catecholamine
granules of the adrenal medulla. Acta Physiol Scand 1959; 45:
328-38.
52. Hingerty D, O'Boyle A. Clinical Chemistry of the Adrenal Medulla.
Springfield, Charles C Thomas, 1972.
53. Holtz P, Shüman HJ. Arternol - Ein neues Hormon des Nebennierenmarks.
Naturwissenschaften 1948; 35: 159.
54. Hopwood D. The histochemistry and electron histochemistry
of chromaffin tissue. Prog Histochem Cytochem 19 7 1; 3: 1-66.
55. Houssay BA, Molinelli EA. Excitabilité des fibres adrenaline-secretories
des nerf grand splanchnique; Fréquences, seu il et optimum
des stimulus role de l'ion calcium. CR Soc Biol 1928; 99: 172-74.
56. Jacobj C. Beiträige zur physiologischen und pharmakologischen
Kenntnis der Darmbewegungen met besonderorer Beruckschtingung
der Beziehung der Nebenniere zu Densel- ben. Arch Exp Path Pharmakol
1892; 29: 171-21 1.
57. Joesten G. Der feinere Bau der Nebennieren. Arch Heilk 97-110.
58. Kirshner N. Uptake
of catecholamines by a particulate fraction of the adrenal medulla.
J Biol Chem 1962; 237: 2311-17.
59. Kohn A. Das chromaffine Gewebe. Ergebnisse Anat Entwicki 1902;
12: 253-348.
60. Kölliker A. Handbuch der Gewebelehre des Menschen. Leipzig:
W Engelmann, 1852.
61. Kölliker A. Manual of Human Histology (translated and
edited by G Busk and T Huxley), London, 1854.
62. LaGamma EF, Adler JE. Glucocorticoids regulate adrenal opiate
peptides. Mol Brain Res 1987; 2: 125-30.
63. Langley JN. On the reaction of cells and of nerve-endings
to certain poisons, chiefly as regards the reaction of striated
muscle to nicotine and to curare. J Physiol (Lond) 1901; 33: 374-413.
64. Le Douarin N, Teillet M. Experimental analysis of the migration
and differentiation of neuroblasts of the autonomic nervous system
and of neuroectodermal mesenchymal derivatives using a biological
cell marking technique. Dev Biol 1974; 41: 162-84.
65. Lenard A The history of research on the adrenals, 1563-1900.
J Hist Med 195 1; 6: 496-505.
66. Lever JD. Electron microscopic observations on the normal
and denervated adrenal medulla of the rat. Endocrinology 1955;
57: 621-35.
67. Lewandowsky M. Ueber die Wirkung des Nebennieren- extracts
auf die glatten Muskeln, irn Besondersen des Auges. Arch Anat
Physiol 1899; 23: 360-66.
68. Lewis RV, Stern AS, Rossier J, Stein S, Udenfriend S. Putative
enkephalin precursors in bovine adrenal medulla. Biochem Biophys
Res Commun 1979; 89: 822-29.
69. Livett BG. Adrenal medullary chromaffin cells in vitro. Physiol
Rev 1984; 64: 1103-61.
70. Madrazo 1, Drucker-Colin R, Diaz V, Martinez-Mata J, Torres
C, Becerril JJ. Open microsurgical autograft of adrenal medulla
to the right caudate nucleus in two patients with intractable
Parkinson's disease. New Eng J Med 1987; 316: 831-34.
71. Manasse P. Über die hyperplastichen Tumoren der Nebennieren.
Virchow Arch Pathol Anat 1893; 133: 391-404.
72. Manasse P. Ueber die Beziehungen der Nebennieren zu den Venen
und dem venosen Kreislauf. Virchow Arch Pathol Anat 1894; 135:
263-76.
73. Manasse P. Zur Histologic und Histogenese der primdren Nierengeschwiiiste.
Virchows Arch Pathol Anat 1896; 145-. 113-57.
74. Manger WM, Gifford RW Jr. Pheochromocytoma. New York: Springer-Verlag,
1977.
75. Mayo CH. Paroxysmal hypertension with tumor of retro-peritoneal
nerve. Report of case. J Am Med Assoc 1927; 89. 1047-50.
76. Meckel JF. Abhandlungen aus der menschilichen und vergleichenden
Anatomic und Physiologic. Halle, 1806.
77. Medvei VC. A History of Endocrinology. Lancaster: MTP Press
Ltd, 1982.
78. Merck Index, 10th edition, M Windhoiz, ed. Rahway, NJ, 1983.
79. Mitchell JS. Obituary, Prof EJ Friedmann. Nature 1956; 178:
397.
80. Nagel (Dr). Ueber die Struktur der Nebennieren. Arch Anat
Physiol Wissen Med. Veriag von G Eichler, Berlin, 1836.
81. Njus D, Knoth J, ZaHakian M. Proton-linked transport in chromaffin
granules. Current Topics Bioenerget 1981; II: 107-47.
82. Njus D, Radda GK. Bioenergetic processes in chromaffin granules.
A new perspective on some old problems. Biochem J 1978; 180: 579-85.
83. Noda M, Furutani Y, Takahashi H, Toyosato M, Hirose T, Inayama
S, Nakanishi S, Numa S. Cloning a sequence analysis of CDNA for
bovine adrenal preproenkephalin. Nature 1982; 295. 202-06.
84. Oliver G, Schäfer EA. On the physiological action of
extract of the suprarenal capsules. J Physiol Lond 1894; 16: i-iv.
85. Pincoffs MC. A case of Paroxysmal hypertension associated
with suprarenal tumor. Trans Assoc Am Physicians 1929; 44. 295-99.
86. Poll H. Verh Physiol Ges 1903; p. 87.
87. Rabin CB. Chromaffin cell tumor of the suprarenal medulla
(pheochromocytoma). Arch Pathol 1929; 7: 228-43.
88. Remak R. Ueber ein selbstaendiges Darrnnervensystem. Berlin,
1847.
89. Rolleston HD. The Endocrine Organs in Health and Disease,
with an Historical Overview. London-Oxford Univ Press, 1936.
90. Schenk J von Grafenberg. Observationum medicarum, rararurn,
novarurn, 2 Vols. Francofurti. JRhodii, 1600.
91. Schmidt JCH. De glandulis suprarenalibus (54 pp), Traj ad
Viadr e typ Winteriano, 1785.
92. Schultzberg M, Lundberg JM, Hökfelt T, Terenius L, Brandt
J, Elde RP, Goldstein M. Enkephalin-like imrnunoreactivity in
gland cells and nerve terminals of the adrenal medulla. Neuroscience
19 78; 3: 1169-86.
93. Sherwin RP. The adrenal medulla, paraganglia and related tissues.
In: Bloodworth JMB Jr, ed, Endocrine Pathology. Baltimore: Williams
&- Wilkins, 1968.
94. Shumacker HB Jr. The early history of the adrenal glands with
particular reference to theories of function. Bull Johns Hopkins
Univ 1936; 4: 39-56.
95. Sietzen M, Schober M, Fischer-Colbrie R, Scherman D, Sperk G, Winkler H. Rat adrenal medulla: Levels of chromogranins, enkephalins, dopamine b-hydroxylase and of the amine transporter are changed by nervous activity and hypophysectomy. Neuroscience 1987 -122: 131-9.
96. Simon M. Sieben Bülcher Anatomie des Galen. JC Hinrichs'schue Buchandlung, Leipzig, 1906.
97. Sjöstrand FS,
Wetzstein R. Elektronenmikroskopische Untersuchchung der phdochromen
(chromaffinen) Granula in den Markzcilen der Nebenniere. Experientia
1956; 12- 196-9.
98. Smith AD, Winkler H. The localization of lysosomal enzymes
in chromaffin tissue. J Physiol (Lond) 1966; 183: 179-88.
99. Smith AD, Winkler H. Purification and properties of an acidic protein from chromaffin granules of bovine adrenal medulla. Biochem J 1967; 103: 483-92.
100. Smith WJ, Kirshner N. A specific soluble protein from the catecholamine storage vesicles of bovine adrenal medulla. Molec Pharrnacol 1967; 3. 52-62.
101. Sorkin SZ. The adrenals before Addison. J Mt Sinai Hosp 1957; 24: 1238-49.
102. Stilling H. Ucber die compensatorische Hypertrophic der Nebennieren. Virchows Arch 1889; 118. 569-75.
103. Szymonowicz L. Die
Function der Nebenniere. Pflueger's Arch Ges Physiol 1896; 64.
97-164.
104. Trifaró J-M, Lee RWH. Morphological characteristics
and stimulus-secretion coupling in bovine adrenal chromaffin cultures.
Neuroscience 1980; 5: 153346.
105. Unsicker K, Krisch B, Otten U, Thoenen H. Nerve growth factor-induced
fiber outgrowth from isolated rat adrenal chromaffin cells - impairment
by glucocorticoids. Proc Nati Acad Sci USA 1978; 75: 3498-502.
106. Valsalva AM. Opera, Dissertatio Anatomica tertia, pp. 151-55,
F. Pitteri, Venetius, 1740.
107. van Heerden JA. First encounters with pheochromocytoma, the
story of Mother Joachim. Am J Surg 1982; 144: 277-79.
108. Virchow R. Arch path Anat 1857; 12. 481.
109. Viveros OH, Diliberto EJ, Hazum E, Chang KJ. Opiate-like
materials in the adrenal medulla: Evidence for storage and secretion
with catecholamines. Mol Pharniacol 1979; 16: 1101-08.
110. von Euler US, Harnberg U. l-Noradrenaline in the suprarenal
medulla. Nature 1949; 163: 642-3.
111. Vulpian M. Note dur quelques reactions propres à la
substance de capsules surrénales. Compt rend Acad Sci (Paris)
1856; 43: 663-65.
112. Welbourn RB. Early surgical history of phaeochrormocytoma.
Brj Surg, 1987; 74: 594-96.
113. Werner B. De Capsulis Suprarenalis, diss ingarg Dorpati Livonorum,
1857.
114. Winkler H. The composition of adrenal chromaffin granule:
An assessment of controversial results. Neuroscience 1976; 1:
65-80.
115. Winkler H. The biogenesis of adrenal chromaffin granules.
Neuroscience 1977; 2.- 657-83.
116. Winkler H. Citation classic. Current Contents 1987; 30: 16.
117. Winkler H, Apps DK,
Fischer-Colbrie R. The molecular function of adrenal chromaffin
granules. Established facts and unresolved topics. Neuroscience
1986; 18: 261-90.
118. Winkler H, Carmichael SW. The chromaffin granule. In: Poisner
and Trifaró, eds, The Secretory Granule. Elsevier Biomed
Press, 1982.
119. Wurtrnan Rj, Axelrod
J. Adrenaline synthesis: Control by the pituitary gland and adrenal
glucocorticoids. Science 1965; 150: 1464-65.
120. Yoburn BC, Franklin SO, Calvano SE, Inturrisi CE. Regulation
of rat adrenal medullary enkephalins by glucocorticoids. Life
Sci 1987; 40: 2495-503.