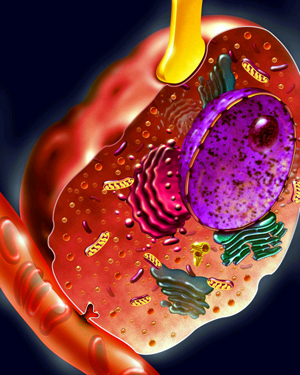
A stylized drawing of a chromaffin cell
is shown above, with a nerve process (in yellow) coming in from
above, and secretory products (adrenaline, etc.) being secreted
into the bloodstream at the lower left of the picture. Chromaffin
cells are found in various parts of the body, but they are most
heavily concentrated in the adrenal medulla. Below is a description
(in English) of chromaffin cells in the adrenal medulla that has
been extracted from a Scientific American article (Carmichael,
S.W. and H. Winkler, The adrenal chromaffin cell, Scientific American
253:40-49, [August] 1985). This article is also available in Spanish
(La célula suprarrenal cromafín, Investigacion y
Ciencia, 109:18-29, 1985), Japanese (Mechanisms of biosynthesis
and secretion in chromaffin cells, Saiens 10:46-58, 1985), German
(Die chromaffinen Zellen der
Nebenniere, Spektrum der Wissenschaft, 10:70-82, 1985), and Italien
( Le cellule cromaffini dei surreni, Le Scienzie, 206:54-65, 1985).
Under conditions of fear or stress, a surge of the hormone adrenaline mobilizes the body for peak physical response. Flooding the bloodstream at up to 300 times the normal concentration, the adrenaline interacts with receptors on cells in various organs, increasing the heart rate and blood pressure and prompting the release from the liver of extra sugar to fuel muscular work. Taken together, these reactions constitute a "fight or flight" response that prepares one to combat an enemy or flee from danger. They are the end result of a secretary event in the adrenal medulla: the inner part of the two adrenal glands just above the kidneys. There, specialized cells known as chromaffin cells manufacture, store, and secrete a complex mixture of hormones, the most important of which is adrenaline.
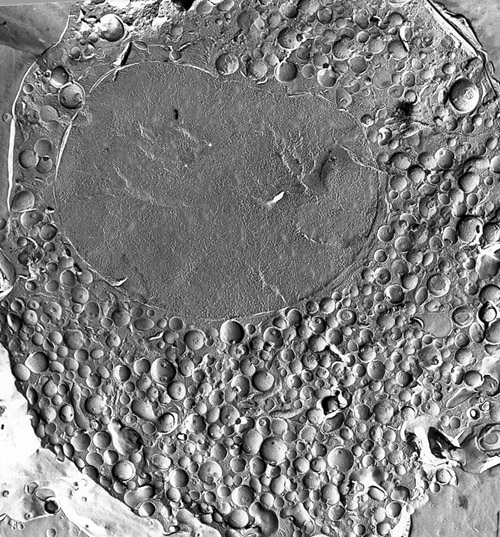
THE CHROMAFFIN CELL encloses a multitude of chromaffin vesicles: intracellular sacs containing adrenaline or noradrenaline and a variety of proteins and peptides. The large oval body is the cell. The subcellular features were highlighted by the freeze-structure technique. An isolated chromaffin cell was quick-frozen and then fractured. Platinum and carbon, sprayed on the fractured surface, formed a replica of the surface relief; the tissue was then dissolved and a transmission electron micrograph was made of the replica. This micrograph was made by Wolfgang Schmidt of the University of Innsbruck.
Chromaffin cells are of interest not only as the root of the fight-or-flight reaction, but also because they offer insights into the workings of other secretory cells, notably neurons, or nerve cells. The adrenal medulla is an endocrine gland: a ductless gland that affects other tissues and organs by discharging hormones into the bloodstream. It can also be thought of, however, as part of the sympathetic nervous system, which helps to regulate such involuntary functions as heart rate, intestinal movements and the dilation of the pupil. Like the neurons of the sympathetic system, the adrenal medulla is controlled by nerves originating in the spinal cord; its primary hormone, adrenaline, is closely related to noradrenaline, the characteristic neurotransmitter of the sympathetic nerves. (Adrenaline and noradrenaline are also known respectively as epinephrine and norepinephrine.) Moreover, the adrenal medulla itself secretes some noradrenaline, and it also releases the neurologically active substances known as neuropeptides.
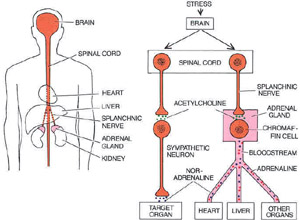 STRESS
LEADS TO SECRETION both in the chromaffin cells of the adrenal
glands, which are just above the kidneys (left), and in
sympathetic neurons, which infiltrate a ]range of tissues and
organs. Impulses arrive at both kinds of cells through nerves
originating in the spinal cord (right). Chromaffin cells
secrete adrenaline, noradrenaline and other substances into the
bloodstream, thereby exerting widespread control over tissues
and organs; sympathetic neurons discharge noradrenaline locally.
Because of the similarities in the activation of chromaffin cells
and sympathetic neurons, in their secretions and in their effects,
and because they develop from the same stem cells (in the neural
crest) during embryogenesis, the adrenal medulla is considered
to be part of the sympathetic nervous system.
STRESS
LEADS TO SECRETION both in the chromaffin cells of the adrenal
glands, which are just above the kidneys (left), and in
sympathetic neurons, which infiltrate a ]range of tissues and
organs. Impulses arrive at both kinds of cells through nerves
originating in the spinal cord (right). Chromaffin cells
secrete adrenaline, noradrenaline and other substances into the
bloodstream, thereby exerting widespread control over tissues
and organs; sympathetic neurons discharge noradrenaline locally.
Because of the similarities in the activation of chromaffin cells
and sympathetic neurons, in their secretions and in their effects,
and because they develop from the same stem cells (in the neural
crest) during embryogenesis, the adrenal medulla is considered
to be part of the sympathetic nervous system.
The secretion of hormones from the chromaffin cells elicits a widespread response. In contrast, neurons, which have fine axons that extend into their target organs, exert localized control at their axon terminals. When chromaffin cells are grown in culture, however, they extend axon-like processes, which indicates their close kinship with neurons. Adrenal chromaffin cells are available in a relatively pure form, unlike sympathetic neurons, which are scattered throughout the body; the cells are therefore accessible to biochemical analysis. As a result they have served as a laboratory model of neurons, and much of what is known about the production and secretion of neurotransmitters was established through studies of the chromaffin cell.
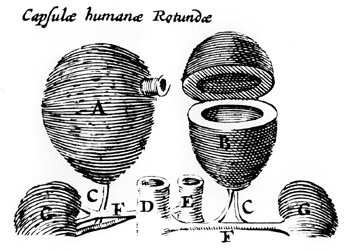 THE
ADRENAL GLANDS (A, B) are shown in a woodcut made in 1611
by the Danish anatomist Caspar Bairtholinus. The medulla of the
adrenal gland, the central part that contains the chromaffin cells,
was not yet recognized as a distinct entity; Bartholinus described
the glands as capsule like organs filled with "black bile."
Two centuries passed before the French anatomist Georges Cuvier
established that the adrenal gland has a solid medulla. It was
later determined that the dark liquid Bartholinus noted results
from the rapid breakdown of the medulla following death. The structures
labeled G in the woodcut are the kidneys, shown disproportionately
small; the other features that are shown are blood vessels.
THE
ADRENAL GLANDS (A, B) are shown in a woodcut made in 1611
by the Danish anatomist Caspar Bairtholinus. The medulla of the
adrenal gland, the central part that contains the chromaffin cells,
was not yet recognized as a distinct entity; Bartholinus described
the glands as capsule like organs filled with "black bile."
Two centuries passed before the French anatomist Georges Cuvier
established that the adrenal gland has a solid medulla. It was
later determined that the dark liquid Bartholinus noted results
from the rapid breakdown of the medulla following death. The structures
labeled G in the woodcut are the kidneys, shown disproportionately
small; the other features that are shown are blood vessels.
Chromaffin cells owe their name to the color produced by a chemical reaction of adrenaline. In the 19th century it was found that an unidentified substance in the adrenal medulla reacts with ferric chloride to produce a greenish color and with chromium salts to produce a yellowish brown. At the beginning of this century Alfred Kohn, working in Prague, coined the term chromaffin for cells stained by chromium. At about the same time adrenaline, the cellular compound responsible for the reaction, was isolated and synthesized in the laboratory.
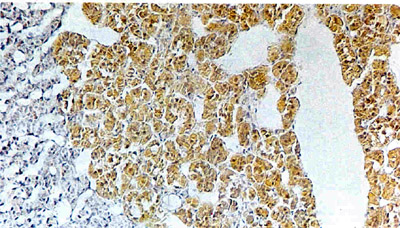 CHROMAFFIN
REACTION yields a yellowish brown color, it takes place when chromium
salts applied to adrenal tissue react with oxidized adrenaline
in the chromaffin cells. In this micrograph of rat adrenal gland
the color distinguishes the adrenal medulla, the site of the chromaffin
cells, from the blue-stained adrenal cortex: the part of the adrenal
gland that surrounds the medulla. The oval space corresponds to
the central vein, which carries off secretions. Rex Coupland of
the University of Nottingham made the image.
CHROMAFFIN
REACTION yields a yellowish brown color, it takes place when chromium
salts applied to adrenal tissue react with oxidized adrenaline
in the chromaffin cells. In this micrograph of rat adrenal gland
the color distinguishes the adrenal medulla, the site of the chromaffin
cells, from the blue-stained adrenal cortex: the part of the adrenal
gland that surrounds the medulla. The oval space corresponds to
the central vein, which carries off secretions. Rex Coupland of
the University of Nottingham made the image.
The first hint of how the chromaffin cell stores adrenaline came in 1953. In that year Hermann Blaschko and Arnold Welch of the University of Oxford and Nils-Åke Hillarp and his coworkers at the Karolinska Institute disrupted cells of the adrenal medulla using special homogenizers and searched for adrenaline and noradrenaline in the resulting mixture. They found the hormones were present not in the cytosol (the watery substance in which the intracellular elements are dispersed) but in a form that settled to the bottom of the tube when the mixture was spun in a centrifuge.
It was apparent that the hormones are stored not as free molecules in the cytosol but within subcellular organelles. Jeffrey Lever, now at University College Cardiff, and other workers made electron micrographs that revealed the organelles. They are small vesicles, or membrane-bounded sacs, about .3 micrometer in diameter. A single chromaffin cell, as calculated by John Phillips of the University of Edinburgh, contains about 30,000 so-called chromaffin granules, or vesicles. The discovery of chromaffin vesicles established for the first time in an endocrine gland a mode of storage now known to be common to almost all cells that secrete hormones or neurotransmitters.
How are the hormones that are contained in chromaffin vesicles synthesized? In 1939, Blaschko proposed that the body makes adrenaline from the amino acid tyrosine in a four-step process, each step being catalyzed by a different enzyme. Since then, other workers have traced the biosynthesis in detail.
The first step, the conversion of tyrosine into dopa, is catalyzed by tyrosine hydroxylase, an enzyme found in the cytosol of chromaffin cells. A second enzyme, dopa decarboxylase, also present in the cytosol, rapidly converts the dopa into dopamine. The enzyme needed to transform dopamine into noradrenaline in the third step of the process, dopamine beta-hydroxylase, is present only within the chromaffin vesicles. Therefore dopamine must be transported into the vesicles for the synthesis to proceed.
Torgeir Flatmark of the University of Bergen and his colleagues have shown the vesicles to be rich in a second substance that is crucial to the production of noradrenaline: ascorbic acid, or vitamin C. Dopamine beta-hydroxylase probably depends on ascorbic acid as a cofactor. The ascorbic acid most likely serves by donating electrons; in the process it is changed to semidehydroascorbate, as Emanuel Diliberto and his colleagues at the Wellcome Research Laboratories have demonstrated. The enzyme needed to reconstitute the ascorbic acid and keep the synthesis of noradrenaline under way is found only in the cytosol, however, necessitating a biochemical link between the vesicle interior and the cytosol. Cytochrome b561, one of the most abundant proteins in the vesicle membrane, is thought to transfer electrons across the membrane to the semidehydroascorbate, thus renewing the supply of ascorbic acid.
In some cells of the adrenal medulla (10 percent of the total in human glands) the process of synthesis ends with noradrenaline, just as it does in the sympathetic nerves. In most adrenal cells, however, noradrenaline is converted into adrenaline by phenylethanolamine N-methyltransferase. Because this enzyme is found only in the cytosol, noradrenaline must leak out of the vesicles to be changed into adrenaline, which is subsequently returned to the vesicles for storage.
Regulatory mechanisms controlling the rate at which adrenaline is synthesized through this four-step process act mainly at the first stage, in which tyrosine is converted into dopa by tyrosine hydroxylase. Both the number of tyrosine hydroxylase molecules and their state of activation determine the rate of synthesis. When the organism is placed under sudden stress, the level of activity of the cell's existing complement of enzyme increases, leading to a spurt in adrenaline production. Norman Weiner and his collaborators at the University of Colorado at Denver School of Medicine showed that in an acutely stressed rat about 50 percent of the tyrosine hydroxylase is in an activated form, whereas only about 5 percent of the enzyme is activated in an unstressed rat.
When stress is sustained, resulting in prolonged stimulation of the adrenal medulla through the splanchnic nerve, a different process leads to a long-term increase in the rate of adrenaline synthesis, Working in the laboratory of Julius Axelrod at the National Institute of Mental Health, Hans Thoenen (now at the Max Planck Institute for Psychiatry in Munich) established that continued stimulation of the chromaffin cell results in the synthesis of extra tyrosine hydroxylase. Coupled with the short-term mechanism of enzyme activation, this long-term adaptive device means the adrenal medulla need not store an excess of hormone sufficient to meet any demand. Similar mechanisms, it is now known, control the synthesis of noradrenaline in sympathetic neurons.
Adrenaline synthesis in the adrenal medulla is also subject to the biochemical influence of the adrenal cortex: the surrounding area of the adrenal gland. Because blood from the cortex passes through the medulla, most of the chromaffin cells are exposed to high levels of cortical steroids, the hormones released by the cortex. When the steroids are absent, Roland Ciaranello and Dona Wong, then at Stanford University School of Medicine, have found the enzyme that catalyzes the conversion of noradrenaline into adrenaline is degraded at an unusually high rate; its level in the cell declines, slowing the synthesis of adrenaline. Thus the intimate anatomical relation of the cortex and the medulla extends to the molecular level as well. Stress is known to induce the secretion of hormones from the adrenal cortex as well as from the medulla. The cortical hormones apparently ensure that the synthesis of adrenaline is maintained.
At two junctures in the process of adrenaline synthesis, crucial substances synthesized in the cytosol of the chromaffin cell must be transported into the chromaffin vesicles. Dopamine must enter the vesicles for conversion into noradrenaline; adrenaline, the end product of the synthesis, must pass from the cytosol into the vesicles for storage. Within the vesicles the concentration of adrenaline is at least 25,000 times as great as it is in the cytosol, testifying to the efficacy of the transport mechanism. How are hormones pumped across the vesicle membrane?
In 1962 Arvid Carlsson and his colleagues at the University of Göteborg and Norman Kirshner of the Duke University School of Medicine reported an early clue to the nature of the transport mechanism. They showed that isolated chromaffin vesicles can accumulate and store adrenaline from a dilute solution. They also discovered that the accumulation took place only when magnesium ions and the compound adenosine triphosphate (ATP), the cellular energy carrier, were included in the solution. Noting that the vesicle membrane contains the enzyme ATPase, which degrades ATP when it is activated by magnesium, Peter Banks, now at the University of Sheffield, proposed that the uptake of adrenaline depends on the energy yielding breakdown of ATP by ATPase. But the actual role of the ATPase in the process remained elusive for 20 years after Banks's proposal.
The role of ATPase became clearer in the late 1970's, when George Radda, David Njus, and their co-workers at Oxford and Robert Johnson and Antonio Scarpa of the University of Pennsylvania School of Medicine demonstrated in an elegant series of experiments that as the ATPase of the vesicle membrane degrades ATP it drives protons (hydrogen ions) from the cytosol into the interior of the vesicles. The accumulation of protons within the vesicle gives its interior a positive charge and makes it more acidic than the cytosol. The difference in charge and acidity across the vesicle membrane is referred to as the electrochemical proton gradient.
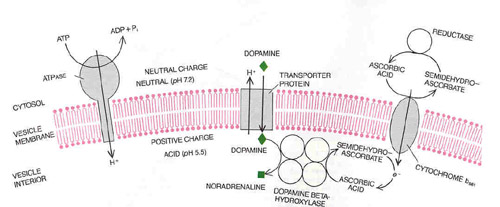
PROTEIN BRIDGES carry catecholamines, protons and electrons across the membrane of the chromaffin vesicle. The enzyme ATPase breaks down ATP in the cytosol into ADP and inorganic phosphate (Pi), releasing energy that pumps protons (H+) across the membrane into the vesicle. A proton gradient is thereby created across the vesicle membrane: the pH of the vesicle interior is lowered, and the vesicle acquires a positive charge. The energy stored in the gradient drives the transport of catecholamines such as dopamine into the vesicle. As protons flow back out into the cytosol a transporter protein carries dopamine into the vesicle. One step in the synthesis of adrenaline, the transformation of dopamine into noradrenaline, takes place inside the vesicle. It is catalyzed by a four-part enzyme, dopamine beta-hydroxylase, with ascorbic acid as a cofactor. In the process the ascorbic acid loses an electron and becomes semidehydroascorbate. A third protein in the vesicle membrane, cytochrome b561, transfers electrons (e-) into the vesicles from a complementary process in the cytosol, thereby renewing the ascorbic acid and enabling the synthesis of noradrenaline to proceed.
The gradient represents stored energy, which drives the uptake of catecholamines (a group that includes adrenaline, noradrenaline and dopamine) as protons flow back down the gradient and out into the cytoplasm. The precise mechanism by which the gradient powers the transport process remains to be determined.
It is clear that to be conveyed into the vesicles the hormones must combine with a special transporter protein. Shimon Schuldiner and Ruth Gabizon of the Hebrew University of Jerusalem recently searched for the protein with a radioactive-labeled molecular probe that was functionally related to the catecholamines. The probe became linked with a vesicle-membrane protein not yet characterized in detail but presumed to be the transporter.
The transport of the catecholamines into the chromaffin vesicles was the first reported instance of a process driven by a proton gradient in an animal-cell organelle other than the mitochondrion. The process in mitochondria was described in 1961 by Peter Mitchell of the Glynn Research Institute in England, who called it the chemiosmotic mechanism. In mitochondria the oxidation of pyruvate (produced by the breakdown of glucose) and other substances generates a proton gradient, which in turn powers the synthesis of ATP. In chromaffin vesicles the relation of ATP and the proton gradient is normally inverted: instead of powering ATP synthesis the proton gradient is sustained by ATP breakdown. Jean-Pierre Henry and Daniel Scherman of the Institute of Physico-Chemical Biology in Paris and Gabriele Taugner of the Max Planck Institute for Medical Research in Heidelberg have shown, however, that when a proton gradient is artificially imposed across the vesicle membrane, the outflow of protons can lead to ATP synthesis.
The ATPase enzymes crucial to the proton gradients of the chromaffin vesicle and of the mitochondrion are made up of distinct protein subunits, as shown by David Apps, working in Edinburgh, and Nathan Nelson and Shulamit Cidon of the Technion-Israel Institute of Technology. The enzymes have a similar two-part structure, however: a component on the membrane surface that takes part in ATP synthesis or breakdown and another component that serves to conduct protons through the membrane.
The uptake mechanism described for catecholamines seems to be common to a number of secretary cells. For example, a proton gradient concentrates serotonin, a substance that is released during blood clotting, in the storage vesicles of blood platelets. A number of investigators have recently established that the accumulation of the neurotransmitter acetylcholine in the synaptic vesicles of neurons also depends on a proton gradient.
Working with our colleagues at the University of Innsbruck, we have demonstrated a second role for the proton gradient in the chromaffin cell. The concentration of ATP is at least 30 times as high in the chromaffin vesicles as it is in the cytosol. Our group has established that the electrochemical gradient, coupled with a distinct transporter protein, carries ATP into the vesicles. Work done by Edward Westhead, Jr., and his colleagues at the University of Massachusetts at Amherst suggests that within the vesicles ATP serves to maintain osmotic stability. Ordinarily a concentrated solution separated from a dilute one by a membrane takes on water by osmosis. Inside chromaffin vesicles the catecholamines form a very concentrated solution; unchecked, osmosis would rupture a chromaffin vesicle. Interaction of the ATP with the catecholamines lowers their effective concentration and thereby averts the process.
The chromaffin vesicles store not only catecholamines and ATP but also a diverse mixture of proteins that may have widespread effects on the nervous system and on other organs. The presence of proteins in the vesicles was recognized in the 1950's by Hillarp; since then the technique of two-dimensional gel electrophoresis, which makes it possible to separate proteins by both their size and their electric charge, has enabled workers to resolve the proteins in the vesicles into at least 30 components. The most abundant component is an acidic protein named chromogranin A.
 PROTEINS
IN THE CHROMAFFIN VESICLE were separated by two-dimensional gel
electrophoresis, in which proteins embedded in a gel are sorted
by electric charge (here expressed as pH) and molecular
size (expressed as molecular weight in thousands). The image at
the left shows the gel after the application of a stain that revealed
the positions of the proteins. Antibodies to specific proteins
were then applied to resolve the vesicle contents into the groups
of proteins shown schematically at the right: chromogranin A
(spots), chromogranins B (outlines), enkephalin
precursors (broken outlines) and the enzyme dopamine beta-hydroxylase.
Reiner Fischer-Colbrie of the University of Innsbruck did the
analysis.
PROTEINS
IN THE CHROMAFFIN VESICLE were separated by two-dimensional gel
electrophoresis, in which proteins embedded in a gel are sorted
by electric charge (here expressed as pH) and molecular
size (expressed as molecular weight in thousands). The image at
the left shows the gel after the application of a stain that revealed
the positions of the proteins. Antibodies to specific proteins
were then applied to resolve the vesicle contents into the groups
of proteins shown schematically at the right: chromogranin A
(spots), chromogranins B (outlines), enkephalin
precursors (broken outlines) and the enzyme dopamine beta-hydroxylase.
Reiner Fischer-Colbrie of the University of Innsbruck did the
analysis.
Although chromogranin A has been isolated and characterized, its function remains obscure and its distribution is still being elucidated. It was originally thought to be confined to the adrenal medulla and the sympathetic nerves. A collaboration between groups led by David Cohn of the Veterans Administration Medical Center in Kansas City and by one of us (Winkler) revealed a similar if not identical protein in the parathyroid gland, however, and chromogranin A has since been found in other endocrine tissues and the brain.
Whatever the other puzzles are that chromogranin A presents, its production within the chromaffin cell is well understood. It is synthesized in the endoplasmic reticulum, the subcellular structure in which many proteins are made, as a proprotein: a precursor of other proteins. It then passes into the Golgi apparatus, the organelle in which intracellular containers such as the chromaffin vesicles are formed, and is packaged into vesicles. Within the chromaffin vesicles the proprotein is split by proteolytic enzymes, freeing smaller molecules.
The features of chromogranin A-its widespread distribution in endocrine and nervous tissues and its proteolytic processing--are common to a group of related but less abundant proteins recently characterized by Reiner Fischer-Colbrie in Innsbruck: chromogranins B. The same features also distinguish the neuropeptides. Neuropeptides are short amino acid chains, some of which act both as neurotransmitters when released by neurons and as hormones when secreted by endocrine glands; they are currently the focus of considerable interest because of their extraordinarily elaborate and varied roles, which may include the regulation of blood pressure, the suppression of pain and the control of behavior [see "Neuropeptides," by Floyd E. Bloom; SCIENTIFIC AMFRICAN, OCTOBER, 1981].
The exciting story of neuropeptides in the adrenal medulla began in 1978, when they were found there by Marianne Schultzberg and Tomas G. M. Hökfelt and their colleagues at the Karolinska Institute. The Karolinska workers exploited immunohistochemistry, in which an antibody to a particular molecule is applied in order to determine the distribution of the molecule in tissues. They showed that the adrenal medulla contains material resembling enkephalins: a group of neuropeptide molecules five amino acids long previously identified in the brain, where they have analgesic properties. Soon afterward 0. Humberto Viveros and his colleagues at the Wellcome Research Laboratories and other groups established that in the adrenal medulla enkephaltins are stored in chromaffin vesicles. While characterizing these enkephalins Sidney Udenfriend of the Roche Institute of Molecular Biology and Jean Rossier of the CNRS Laboratory for Nervous Physiology at Gif-sur-Yvette and their colleagues also found longer peptides that seem to be the precursors of enkephalins.
 PREPROENKEPHALIN
MESSAGE in cultured bovine chromaffin cells that were stiinul2ted
artifici2ily was made visible by Ruth Siegel of the National Institute
of Mental Health by means of in situ hybridization histochemistry.
A sequence of DNA complementary to the messenger RNA (mRNA) coding
for preproenkephalin was prepared and labeled with a radioactive
isotope. When the complementary DNA (cDNA) was introduced into
chromaffin cells, it hybridized with (bound to) the mRNA. The
cells were then coated with a photographic emulsion to make an
autoradiograph, in which black dots reveal the location of radioactive
cDNA that is bound to the mRNA. (Some dots are visible outside
cells because the radioactive particles traveled beyond the cell
before interacting with the emulsion.) The image shows about three
times as many dots as images of unstimulated cells show. Such
results suggest that the increase in the synthesis of preproenkephalin
that occurs in stimulated chromaffin cells stems from an increase
in mRNA encoding the protein.
PREPROENKEPHALIN
MESSAGE in cultured bovine chromaffin cells that were stiinul2ted
artifici2ily was made visible by Ruth Siegel of the National Institute
of Mental Health by means of in situ hybridization histochemistry.
A sequence of DNA complementary to the messenger RNA (mRNA) coding
for preproenkephalin was prepared and labeled with a radioactive
isotope. When the complementary DNA (cDNA) was introduced into
chromaffin cells, it hybridized with (bound to) the mRNA. The
cells were then coated with a photographic emulsion to make an
autoradiograph, in which black dots reveal the location of radioactive
cDNA that is bound to the mRNA. (Some dots are visible outside
cells because the radioactive particles traveled beyond the cell
before interacting with the emulsion.) The image shows about three
times as many dots as images of unstimulated cells show. Such
results suggest that the increase in the synthesis of preproenkephalin
that occurs in stimulated chromaffin cells stems from an increase
in mRNA encoding the protein.
Subsequent research showed how enkephalins are produced in the chromaffin cell. Using the techniques of molecular cloning, Michael Comb of the University of Oregon, Ueli Gubler of the Roche Institute and Masaharu Noda and Shosaku Numa of Kyoto University found that in the adrenal medulla the initial protein, called preproenkephalin, consists of 267 amino acids and contains seven enkephalin sequences. The protein enters the endoplasmic reticulum as it is synthesized; there it is transformed into a proenkephalin through the removal of an initial signal peptide (which enabled the protein to enter the reticulum). The proenkephalin then moves into the Golgi region and is enclosed in chromaffin vesicles, where proteolytic enzymes process it to yield free enkephalins.
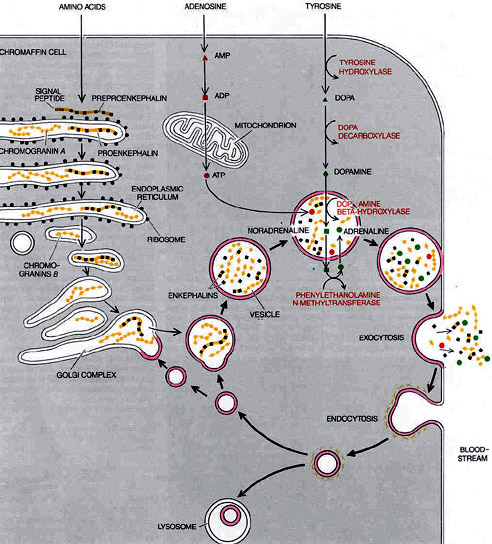
PATHWAYS IN THE CHROMAFFIN CELL include the biochemical routes by which the cell synthesizes the contents of the chromaffin vesicles and the physical cycle through which each vesicle is emptied and its membrane retrieved for subsequent reuse. The primary secretary product of most chromaffin cells, adrenaline, is synthesized from tyrosine in four steps, each one catalyzed by a different enzyme. Tyrosine is transported into the cell, changed into dopa and then into dopamine. The dopamine is carried into the chromaffin vesicles and there is transformed into noradrenaline, which subsequently leaks out of the vesicles into the cytosol and is converted into adrenaline; the adrenaline is pumped back into vesicles for storage. A second substance stored in the vesicles, adenosine triphosphate (ATP), is synthesized from adenosine, which is converted first into adenosine monophosphate (AMP) and then into adenosine diphosphate (ADP). The ADP enters the mitochondrion . and is changed into ATP. Amino acids serve as raw material for the manufacture of enkephalin precursors and chromogranins A and B. Proenkephalin, a long polypeptide chain containing enkephalin sequences, is synthesized on ribosomes along the endoplasmic reticulum. As it passes into the reticulum the preproenkephalin loses a short signal peptide; the shortened molecule (now known as proeukephalin) then moves into the Golgi complex and is packaged in newly formed chromaffin vesicles. There it is broken down into smaller pieces, including some free enkephalins. The chromogranins are also synthesized as large proproteins, most of which are then broken down. The vesicles themselves go through 2 cycle of exocytosis and endocytosis (heavy black arrows). In exocytosis they the move to the cell surface, fuse with the plasma membrane and discharge their contents. The vesicle membrane then takes on a "coated," or fuzzy, appearance, a sign of imminent endocytosis. It pinches off into an endocytotic vesicle and returns to the Golgi region, where the membrane is recycled to make new chromaffin vesicles. Some of the retrieved vesicle membrane does not complete the cycle but is broken down in digestive organelles known as lysosomes.
Like the pace of catecholamine synthesis, the rate at which chromaffin cells produce enkephalins has been found to vary. When chromaffin cells isolated from the adrenal gland of a cow are artificially stimulated, the measured level of enkephalin activity increases along with that of the catecholamines. Lee Eiden and Erminio Costa and their colleagues, working separately at the National Institute of Mental Health, have shown that stimulation increases the amount of messenger RNA coding for preproenkephalin. In the rat stimulation of the chromaffin cell has the opposite effect: Ira Black and his colleagues at Cornell University have demonstrated that it dramatically decreases the enkephalin content even as the synthesis of catecholamines rises.
Although
much is known about the synthesis of neuropeptides in the adrenal
gland and, by extension, in other tissues, we are far from understanding
how these substances function after they are released. James Lewis
and John Liebeskind and their colleagues at the University of
California at Los Angeles have shown that the secreted products
of the adrenal medulla may provide some analgesia during stress
--an effect for which the enkephalins are probably responsible.
On the whole, however, the presence of neuropeptides of various
sizes in the "cocktail" of substances stored in the
adrenal medulla remains mysterious. Insight into why the adrenal
medulla mixes such an elaborate cocktail will eventually help
to explain the secretion of similar mixtures by other endocrine
glands and by nerves. ELECTRON ENERGY-LOSS SPECTROSCOPY reveals
the relative abundances of elements in a chromaffin vesicle and
the surrounding cytosol. The technique uses variations in the
energy loss of the electrons passing through a specimen under
an electron microscope to map the distribution of elements in
the tissue. In this computer-generated image nitrogen is represented
as pink and carbon as blue. The suffusion of pink in the vesicle
reflects its high concentration of nitrogen-containing substances
such as adrenaline, noradrenaline, dopamine and ATP. The blue
halo around the vesicle probably results from the relatively high
carbon content of its membrane. Richard Ornberg and Richard Leapman
of the National Institutes of Health made this image.
ELECTRON ENERGY-LOSS SPECTROSCOPY reveals
the relative abundances of elements in a chromaffin vesicle and
the surrounding cytosol. The technique uses variations in the
energy loss of the electrons passing through a specimen under
an electron microscope to map the distribution of elements in
the tissue. In this computer-generated image nitrogen is represented
as pink and carbon as blue. The suffusion of pink in the vesicle
reflects its high concentration of nitrogen-containing substances
such as adrenaline, noradrenaline, dopamine and ATP. The blue
halo around the vesicle probably results from the relatively high
carbon content of its membrane. Richard Ornberg and Richard Leapman
of the National Institutes of Health made this image.
The chromaffin cell not only stores but also releases its characteristic substances as a mixture. The finding in the 1960's that the adrenal medulla secretes chromogranins and other large molecules at the same time as it releases adrenaline suggested the secretion occurs by exocytosis. In exocytosis a vesicle moves toward the surface of the cell, the vesicle membrane fuses with the plasma membrane (the cell's outer membrane) and the vesicle is opened to the exterior of the cell. The entire content of the vesicle is thereby emptied into the extracellular space.
The first convincing morphological evidence that exocytosis is responsible for secretion from chromaffin cells came in 1967 in the form of electron micrographs of the hamster adrenal gland made by Odile Grynszpan-Winograd (née Diner) of the University of Paris. The micrographs showed chromaffin vesicles opening directly into the extracellular space after fusing with the plasma membrane. Since then exocytosis has been established as the primary release mechanism for all neurotransmitters and hormones, except for the steroids secreted by the adrenal cortex and the gonads.
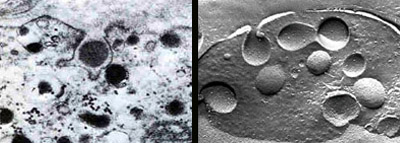
EXOCYTOSIS takes place as the membrane of a chromaffin vesicle fuses with the cell's surface membrane, thus opening the vesicle to the exterior of the cell and releasing its cargo of hormones and other substances. The exocytosis of a single vesicle is shown at a magnification of 40,000 diameters in both a conventional electron micrograph (left), made by Odile Grynszpan-Winograd, and a freeze-fracture micrograph (right), by Wolfgang Schmidt.
Exocytosis from vesicles is an elegant and efficient mechanism of release. It delivers compounds to the exterior of the cell at a much higher concentration than would be possible if they were secreted from the cytosol. It renders unnecessary a specialized mechanism for transporting large molecules across the plasma membrane, which would be needed if secretion took place through an intact membrane. And it enables a cell to release a package of diverse substances as readily as if its membrane were ruptured, but without a loss of cytosol. One of us (Winkler) recently calculated that when a vesicle fuses with the plasma membrane in the chromaffin cell of the cow, it releases about three million molecules of catecholamines, 800,000 molecules of nucleotide compounds such as ATP, 5,000 molecules of chromogranin A, 80 molecules of chromogranins B and several thousand molecules of enkephalin precursors and free enkephalins.
How is exocytosis triggered in the chromaffin cell? When a nerve impulse causes the neurons innervating the adrenal medulla to fire, they release the neurotransmitter acetylcholine. The acetylcholine interacts with receptors on the chromaffin cells, setting in motion a process in which calcium channels (molecular pores in the cell membrane) open and allow the concentration of calcium ions within the cells to increase. Secretion ensues.
Just how an increase in intracellular calcium leads to secretion by exocytosis is still not known. One step toward establishing the role of calcium was to separate the events associated with the influx of calcium ions from the effects of calcium within chromaffin cells. The step became possible when Bruce Livett, now at the University of Melbourne, and his colleagues pioneered techniques for isolating chromaffin cells. Piercing the membrane of the cell makes it possible to change the intracellular calcium level with out opening membrane channels. Peter Baker and Derek Knight of King's College in London caused a selective breakdown of the membrane with brief, intense electric fields; Jack Brooks of Marquette University, Ronald Holz of the University of Michigan Medical School and others found that somewhat larger holes can be made using detergents. Because the pierced membrane is permeable to ions, the addition of calcium to the fluid bathing the cells leads directly to an increase in intracellular calcium.
These techniques enabled investigators to determine that not only calcium but also ATP and magnesium had to be present in the bathing solution for exocytosis to take place. It seems likely that a magnesium-activated ATPase breaks down ATP to provide energy for exocytosis. It is clear as well that calcium's effect is exerted w4thin the cell. What does it interact with there to precipitate exocytosis?
Like most cells, chromaffin cells have a cytoskeleton: an internal network of microtubules and microfilaments made up of structural and contractile proteins. For a while it was thought that in the presence of calcium and possibly ATP the mesh of proteins pulls the chromaffin vesicles toward the edge of the cell, enabling them to fuse with the plasma membrane. This scenario now appears unlikely. Dominique Aunis and his colleagues at the National Institute for Health and Medical Research in Strasbourg have suggested recently that in the presence of calcium the cytoskeletal mesh simply releases the vesicles, allowing them to move toward the plasma membrane. In a related finding Velia Fowler and Harvey Pollard of the National Institute of Arthritis, Diabetes, and Digestive and Kidney Diseases showed that the calcium decreases the viscosity of solutions containing vesicles and contractile proteins such as those of the cytoskeleton. The necessary concentration of calcium was comparable to the level measured in the chromaffin cell during secretion.
Once the vesicles reach the surface of the cell, other proteins presumably mediate the fusion of the vesicles to the plasma membrane. Pollard and Carl Creutz and their colleagues noted that chromaffin vesicles isolated from the cell fuse when they are exposed to calcium and proteins from the cytoplasm. They isolated one of the proteins, which was particularly effective at promoting fusion, and called it synexin, from a Greek word for "meeting." Other synexins have since been identified, but the role of these proteins in exocytosis is uncertain: the fusion of chromaffin vesicles in vitro may not be a valid model for the fusion of vesicles to the plasma membrane in vivo. Some other proteins, which bind to chromaffin vesicles if calcium is present, have been identified and called chromobindins. Their significance for exocytosis is similarly unclear.
Immunologic techniques may establish the role of these or other proteins in the process of exocytosis. One introduces an antibody into a chromaffin cell, where it binds to, and thus neutralizes, a specific protein. By then determining the point at which exocytosis is interrupted it should be possible to define the precise function of the neutralized protein. Labeling the antibody with a fluorescent dye or small gold particles, which are visible under the electron microscope, should make it possible to trace the protein's distribution within the chromaffin cell.
Introducing such an immunologic probe into the chromaffin cell is not simple. One solution was devised by Jose Trifaró and his colleagues at McGill University; their results emphasize the importance of calcium in exocytosis. They ruptured red blood cells and allowed them to re-form in a medium containing antibody to calmodulin, a protein that is crucial to the action of calcium ions within cells. The red cells were in effect transformed into cellular containers of antibody. The workers then introduced the anticalmodulin into chromaffin cells by fusing them with the red cells. The result was an inhibition of secretion from the chromaffin cells, indicating a direct role for calmodulin, and therefore for calcium, in exocytosis.
The release of a hormone or a neurotransrnitter by exocytosis would seem to present a mechanical problem. When a vesicle discharges its contents, its membrane is incorporated into the cell-surface membrane. As secretion proceeds and vesicle membranes are added to the cell surface, the cell might be expected to enlarge to an untenable size. Yet chromaffin cells stay much the same size during secretion.
It was long suspected that some of the surplus surface membrane is returned to the interior of the cell. Eric Holtzman and his colleagues at Columbia University and William Douglas and his colleagues at the Yale University School of Medicine demonstrated that the retrieval takes place. They introduced tracer molecules into the medium surrounding chromaffin cells and later detected the tracers in the cell interior. The workers concluded that chromaffin cells remove surplus membrane by endocytosis. Endocytosis reverses the sequence of exocytosis: part of the surface membrane forms a pocket, pinches off into an endocytotic vesicle and passes into the cell interior, carrying with it some of the extracellular medium.
Since then Erwin Neher and Alain Marty at the Max Planck Institute for Biophysical Chemistry in Göttingen and their colleagues have adapted the electrophysiological technique of patch clamping to detect individual events of endocytosis as well as exocytosis. A micropipette filled with an electrically conducting fluid is attached to a cell, forming a tight electrical seal. By this means electrical events occurring in the patch of cell membrane encircled by the pipette can be measured; the patch of membrane can also be ruptured, allowing electrical events in the rest of the cell membrane to be recorded. The technique reveals minute changes (on the order of 10 nano farad) in the electrical capacitance of the plasma membrane. Under certain conditions such electrical events may be associated with the addition or removal of a small piece of membrane, such as that of a chromaffin vesicle.
Two questions remained: Does endocytosis single out what had been a vesicle membrane for retrieval instead of simply absorbing an equivalent patch of plasma membrane? If it does, how long does the cycle of exocytosis and subsequent endocytosis take? To answer these questions several groups, including ours in Innsbruck, exposed chromaffin cells to antibodies against proteins known to be unique to the inner surface of the vesicle membrane. The antibodies were labeled with a fluorescent dye, making it possible to identify the membrane following exocytosis. When isolated chromaffin cells were stimulated, the labeled vesicle membranes became visible as discrete spots on the plasma membrane, indicating that exocytosis had taken place. After the stimulation was stopped the fluorescent patches corresponding to vesicle membranes disappeared from the cell surface, having been internalized by endocytosis; the retrieval was complete within 30 minutes.
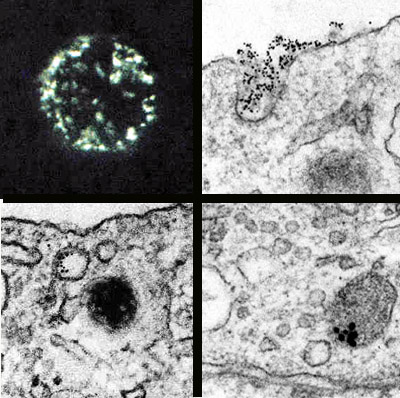
MEMBRANE RECYCLING is documented in micrographs made by Alexandra Patzak of the University of Innsbruck. The photomicrograph (top left) shows the surface membrane, of a cell that was stimulated to secrete. The cell was treated with a fluorescent antibody to a protein specific to the inner surface of chromaffin vesicles. Patches of the membrane give off a green glow, showing that they are bits of vesicle membrane that have been incorporated into the surface membrane through exocytosis. Labeling the antibody with electron-dense colloidal gold enables one to follow the recycling of such patches of membrane in electron micrographs. First the vesicle membrane, decorated with gold-labeled antibodies (visible as black dots), takes on a coated appearance and forms a pocket (top right), which will later become an endocytotic vesicle. Within such a vesicle labeled membrane is visible (bottom left). A micrograph made in a different experiment records the completion of the cycle: former vesicle membrane is evident in an organelle that appears to be a new chromaffin vesicle (bottom right).
To determine the fate of the vesicle membranes after retrieval we needed an immunologic probe that could be resolved at the level of individual organelles, under the electron microscope. In place of fluorescent antibodies we used antibodies tagged with electron-dense gold particles to mark vesicle membranes during their exposure on the cell' surface. We then followed the retrieval of the membranes in a series of electron micrographs. The labeled patches first took on a fuzzy aspect characteristic of "coated pits": patches of cell membrane about to undergo endocytosis. The gold-labeled coated pits then formed pockets and pinched off to become endocytotic vesicles within the cell. In time the vesicle membranes lost their coating. Some of them could later be seen in the Golgi apparatus, where new vesicles are formed. In the end we found gold-labeled vesicle membrane in what appeared to be newly formed vesicles.
Earlier studies of several endocrine glands by Marilyn Farquhar of Yale, Volker Herzog of the University of Munich and Berton Pressman and Robert Rubin of the University of Miami School of Medicine used nonspecific markers to demonstrate that membrane is recycled between the Golgi region and the cell surface. The use of specific antibodies has now enabled us to establish that in the chromaffin cell the membrane of the secretory vesicle is recycled through vesicle formation, exocytosis and endocytosis. Is it not an elegant and economical strategy to use the vesicle membrane as a recyclable container?
The adrenal chromaffin cell has demonstrated its worth as a laboratory model for neurons and other secretory cells. The chromaffin cell may soon prove itself as a substitute for neurons in a clinical setting as well. In 1982 Lars Olson and his colleagues at the Karolinska Institute did the first neural transplants performed in human beings: they transferred chromaffin cells from the adrenal medulla to the brain in patients with severe Parkinson's disease. In Parkinsonism the dopamine-containing neurons in a certain area of the brain stem degenerate. From a wealth of basic research the workers reasoned that chromaffin cells, which produce dopamine as a precursor to noradrenaline, could compensate for the dopamine deficiency.
Of the two patients treated with the experimental surgery, one showed significant improvement and the other did not get worse. The possibility that the adrenal chromaffin cell will one day figure in the treatment of Parkinson's disease typifies the way basic biological research, done for the mere joy of discovery, can yield unexpected benefits for humanity.
STEPHEN CARMICHAEL and HANS WINKLER began their intercontinental collaboration on the adrenal medulla during Carmichael's 1979 sabbatical leave in Innsbruck. Carmichael is a Professor and Chair of Anatomy and Professor of Orthopedic Surgery at the Mayo Clinic, where he teaches gross anatomy, cytology, histology and embryology to medical students and residents. He was graduated in 1967 from Kenyon College and in 1971 from Tulane University with a Ph.D. in Anatomy. Winkler is Chairman of the Department of Pharmacology at the University of Innsbruck, where he got his M.D. in 1963. He also has a D.Phil. from the University of Oxford, which he earned in 1968 for work done in the laboratory of Hermann Blaschko. He became Professor of Pharmacology at Innsbruck in 1975, served as Dean of the Faculty of Medicine from 1977 to 1979 and was made Department Chairman in 1980.
THE ADRENAL MEDULLA. Stephen W. Carmichael. Eden Press, 1979-84.
THE MOLECULAR ORRANIZATION OF ADRENAL CHROMAFFIN GRANULES.
Hans Winkler and Edward Westhead in Neuroscience, Vol. 5, No. I 1, pages 1803-1823; November, 1980.
THE CHROMAFFIN GRANULE. H. Winkler and S. W. Carmichael in The Secretory Granule, edited by A. M. Poisner and J. M. Trifaró. Elsevier Biomedical Press, 1982.
CALCIUM CONTROLS EXOCYTOSIS IN BOVINE ADRENAL MEDULLARY CELLS.
Peter Baker and Derek Knight in Trends in Neuroscience, Vol. 7, No. 4, pages 120-126; April, 1984.